
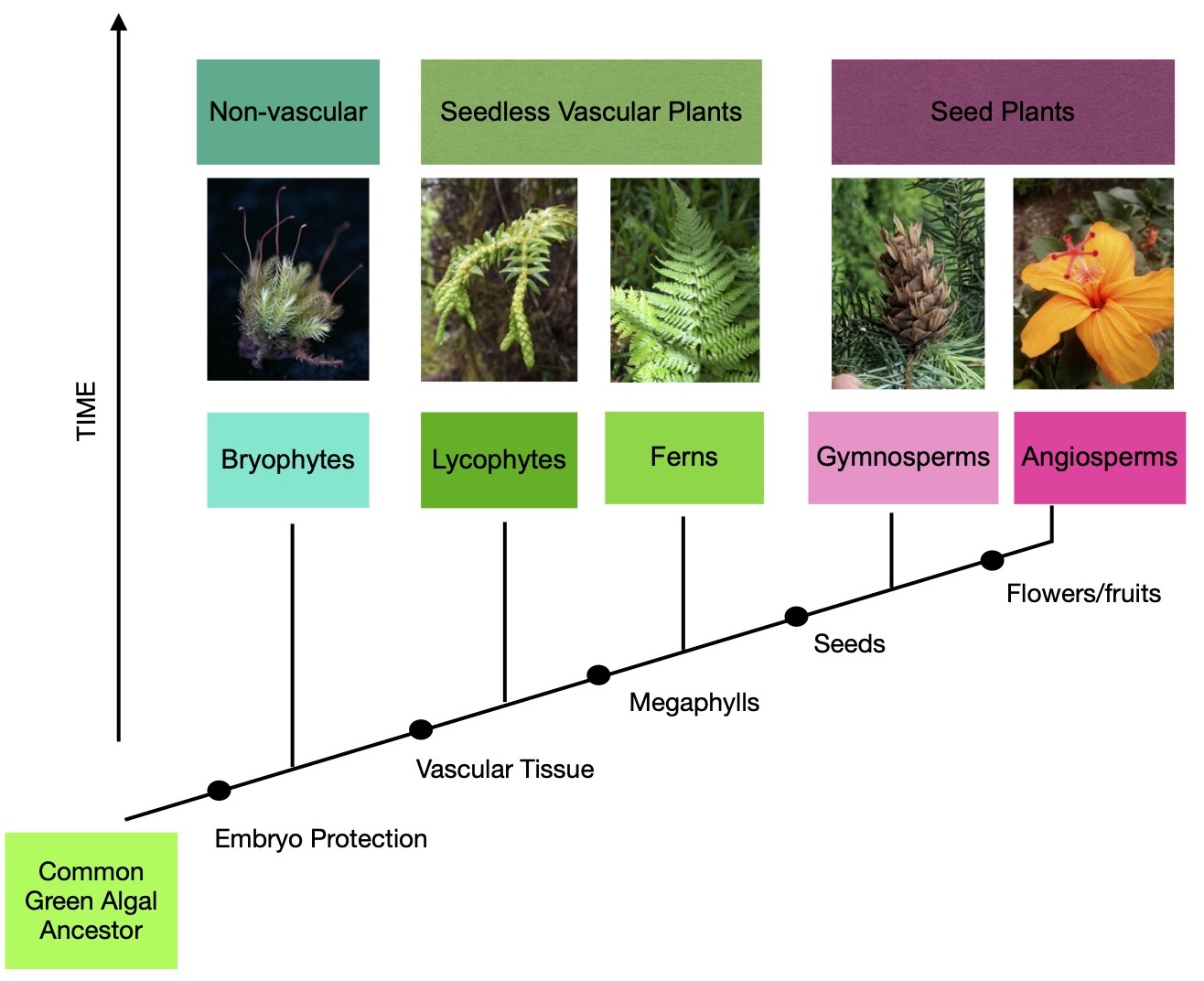
9.1: Angiosperms
- Page ID
- 59250

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Angiosperms are the group of plants that we are most familiar with, as most of the fruits and vegetables we eat belong to this group. They are currently the dominant group of plants in most terrestrial ecosystems and are easy to recognize since they produce flowers (Figure \(\PageIndex{1}\)). Angiosperms are the most diverse group of plants, with 416 families and approximately 352,000 species (APG, 2016). The first fossils of angiosperms date back to the Lower Cretaceous (~130 million years ago; Magallón et al., 2013; Appendix 1). Over time, they diversified and became more prominent in terrestrial ecosystems, slowly displacing and replacing gymnosperms and other plants. Today, angiosperms represent 90% of terrestrial plants on the planet. The remaining terrestrial plant species comprise gymnosperms (approximately 1% of species; Crepet and Niklas, 2009) and bryophytes, lycophytes, and ferns (9% of species).
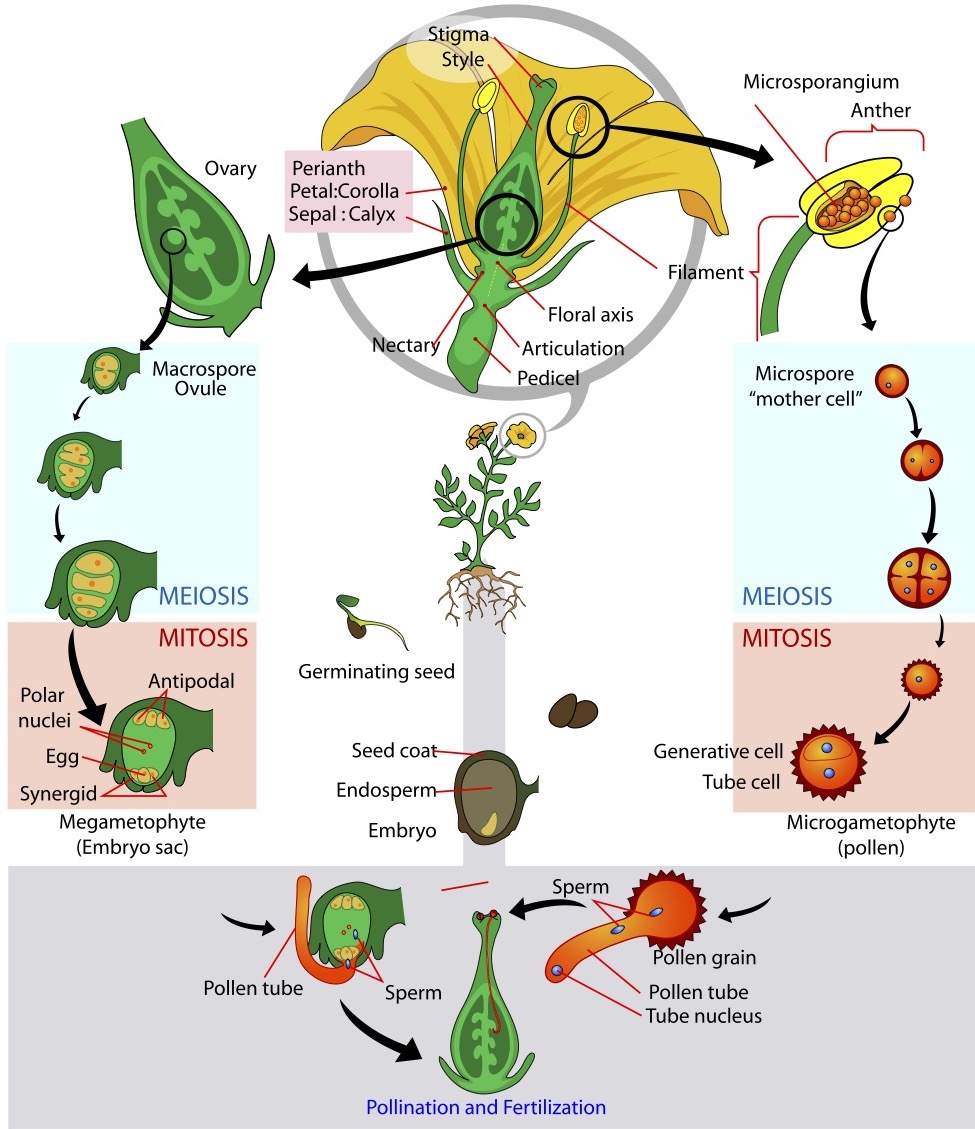
The diversity and success of angiosperms can be attributed to the evolution of flowers and fruits within this group. The development of flowers was an innovative characteristic, as it allowed the ovules to be enclosed and protected inside an ovary, which after fertilization turns into a fruit that aids seed dispersal. Flowers also allowed for male and female parts of the plant to be clustered into a single structure (Figure \(\PageIndex{2}\)).
Flowering plants developed relationships with pollinators (Hu et al., 2008). Instead of dispersing their pollen via wind, like gymnosperms, early angiosperms started to offer rewards in their flowers to attract animals, who in turn started to visit the flowers of a given species for the food rewards they offered. Through this relationship, flowering plants got the guarantee that their pollen would likely reach another individual of the same species: therefore, increasing the probability of successful fertilization (Figure \(\PageIndex{3}\)). This transfer of pollen from one plant to another, otherwise known as cross-pollination, ensured higher genetic diversity in the population.

Since the origin of flowers, pollinators have driven floral diversity in angiosperms by increasing the richness of species (Van der Niet and Johnson, 2012). Pollinators have preferences for flower shape, size, color, and even type of reward. They also come in all shapes and sizes and they include birds, reptiles, beetles, and bats. Over time, some plants and pollinators have developed a specific relationship, where a plant species is pollinated by a specific animal. However, many plants (and animals) are generalists and are pollinated by different species of pollinators.
The Hawaiian honeycreepers are a great example of the relationship between pollinators and flowering plants. At least five genera of a group of plants called Hawaiian lobeliads (Delissea, Clermontia, Cyanea, Lobelia and Trematolobelia) are pollinated by native forest birds (Pender et al., 2014). The shape of the flowers, the nectar content, and other characteristics match this kind of pollination syndrome. The native Hawaiian honeycreeper ‘i'iwi (Vestiaria coccinea) feeds on the nectar produced by these flowers thanks to its long, curved beak which has a similar shape to the curved flowers of many of the Hawaiian lobeliads (Figure \(\PageIndex{4}\)). When feeding on nectar, pollen from the flower is deposited on the head of the ‘i'iwi, and the bird inadvertently moves pollen between flowers. Unfortunately, it is believed that many Hawaiian lobeliad species produce reduced amounts of seed because some native bird species are so rare or have become extinct; therefore, the flowers are not adequately pollinated by these native birds.

A long, tubular Hawaiian lobeliad flower could not be pollinated by insects like bees because of its curved shape. On the other hand, some plant species like ʻōhiʻa (Metrosideros polymorpha) are known as generalists, as they can be pollinated by either birds or insects (Figure \(\PageIndex{5}\)). The flower shape and other characteristics, like color, allow for this type of pollination strategy.

Fruits are another evolutionary adaptation present in angiosperms. Once the plants are pollinated and the ovules fertilized, the ovary grows into a fruit. One of the main functions of the fruit is to assist in the dispersal of the seeds, which is achieved through different mechanisms. For example, fruits can be attractive to animals because they offer a food reward. After being eaten, the seeds contained in the fruit are usually dispersed a distance from the parent plant, allowing the species to spread. Other fruits have spurs to attach themselves to animal fur and bird feathers to aid the dispersal of seeds. An example of the importance of animals in the dispersal of angiosperm seeds is exemplified by the long-distance dispersal of seeds to Hawai‘i from distant land masses; bird dispersal is the principal mode in which native plant genera arrived in the islands.