
6.5: Non-homologous end joining leads to translocations
- Page ID
- 143820
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Repeated sequences are not only responsible for ectopic recombination. In combination with DNA damage, they can also lead to chromosome translocations, where part of a chromosome breaks off and is reattached to a different chromosome, often with disastrous consequences -- they are particularly prevalent in cancer cells.
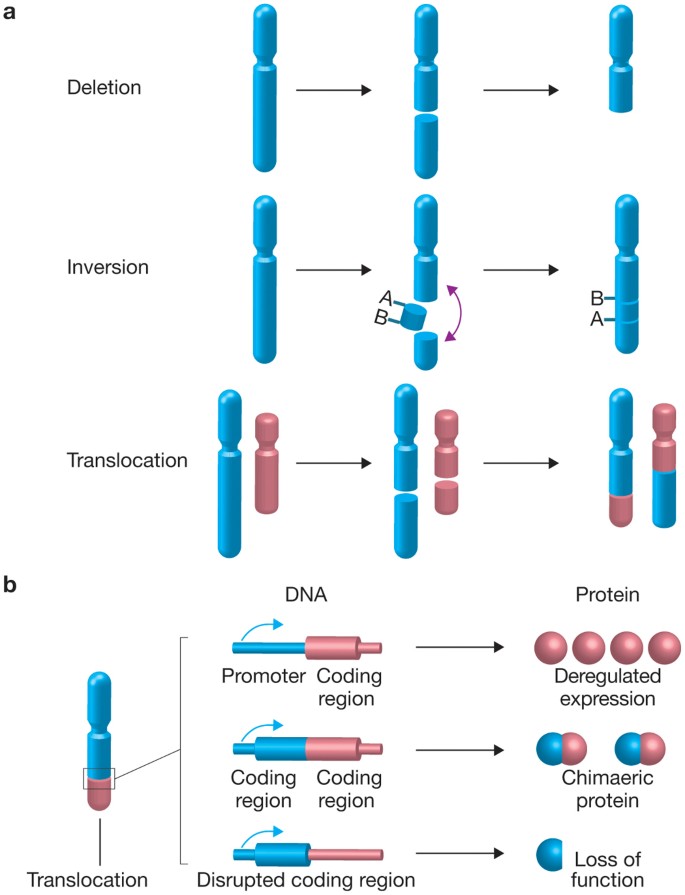
(credit: Roukos and Misteli, The biogenesis of chromosome translocations, Nature Cell Biology 16:293 (2014). https://www.nature.com/articles/ncb2941)
Translocations arise when a DNA molecule sustains damage and breaks entirely. The cell then attempts to repair the break using non-homologous end joining or NHEJ.
Non-homologous end joining
Non-homologous end joining (NHEJ) is a pathway that repairs double-strand breaks in DNA. It is called "non-homologous" because the break ends are directly ligated without the need for a homologous template, in contrast to homology directed repair (HDR), which requires a homologous sequence to guide repair. NHEJ is active in both non-dividing and proliferating cells, while HDR is not readily accessible in non-dividing cells. The term "non-homologous end joining" was coined in 1996 by Moore and Haber.
NHEJ is typically guided by short homologous DNA sequences called microhomologies. These microhomologies are often present in single-stranded overhangs on the ends of double-strand breaks. When the overhangs are perfectly compatible, NHEJ usually repairs the break accurately. Imprecise repair leading to loss of nucleotides can also occur, but is much more common when the overhangs are not compatible. Inappropriate NHEJ can lead to translocations and telomere fusion, hallmarks of tumor cells.
Chromosomal translocation
In genetics, chromosome translocation is a phenomenon that results in unusual rearrangement of chromosomes. This includes balanced and unbalanced translocation, with two main types: reciprocal, and Robertsonian translocation. Reciprocal translocation is a chromosome abnormality caused by exchange of parts between non-homologous chromosomes. Two detached fragments of two different chromosomes are switched. Robertsonian translocation occurs when two non-homologous chromosomes get attached, meaning that given two healthy pairs of chromosomes, one of each pair "sticks" and blends together homogeneously.
A gene fusion may be created when the translocation joins two otherwise-separated genes. It is detected on cytogenetics or a karyotype of affected cells. Translocations can be balanced (in an even exchange of material with no genetic information extra or missing, and ideally full functionality) or unbalanced (where the exchange of chromosome material is unequal resulting in extra or missing genes).
Reciprocal translocations
Reciprocal translocations are usually an exchange of material between non-homologous chromosomes and occur in about 1 in 491 live births. Such translocations are usually harmless, as they do not result in a gain or loss of genetic material, though they may be detected in prenatal diagnosis. However, carriers of balanced reciprocal translocations may create gametes with unbalanced chromosome translocations during meiotic chromosomal segregation. This can lead to infertility, miscarriages or children with abnormalities. Genetic counseling and genetic testing are often offered to families that may carry a translocation. Most balanced translocation carriers are healthy and do not have any symptoms.

Courtesy: National Human Genome Research Institute - [1] (file), Public Domain, https://commons.wikimedia.org/w/inde...p?curid=935175
It is important to distinguish between chromosomal translocations that occur in germ cells, due to errors in meiosis (i.e. during gametogenesis), and those that occur in somatic cells, due to errors in mitosis. The former results in a chromosomal abnormality featured in all cells of the offspring, as in translocation carriers. Somatic translocations, on the other hand, result in abnormalities featured only in the affected cell and its progenitors, as in chronic myelogenous leukemia with the Philadelphia chromosome translocation.
Nonreciprocal translocation
Nonreciprocal translocation involves the one-way transfer of genes from one chromosome to another nonhomologous chromosome.
Robertsonian translocations
Robertsonian translocation is a type of translocation caused by breaks at or near the centromeres of two acrocentric chromosomes. The reciprocal exchange of parts gives rise to one large metacentric chromosome and one extremely small chromosome that may be lost from the organism with little effect because it contains few genes. The resulting karyotype in humans leaves only 45 chromosomes, since two chromosomes have fused together. This has no direct effect on the phenotype, since the only genes on the short arms of acrocentrics are common to all of them and are present in variable copy number (nucleolar organiser genes).

Original from DataBase Center for Life Science (DBCLS)Derivative by Mikael Häggström, M.D. Author info- Reusing images- Conflicts of interest:NoneMikael Häggström, M.D. - File:202208 Chromosomal structural abnormality robertsonian translocation icon.svg, CC BY 4.0, https://commons.wikimedia.org/w/inde...urid=124442249
Robertsonian translocations have been seen involving all combinations of acrocentric chromosomes. The most common translocation in humans involves chromosomes 13 and 14 and is seen in about 0.97 / 1000 newborns. Carriers of Robertsonian translocations are not associated with any phenotypic abnormalities, but there is a risk of unbalanced gametes that lead to miscarriages or abnormal offspring. For example, carriers of Robertsonian translocations involving chromosome 21 have a higher risk of having a child with Down syndrome. This is known as a 'translocation Downs'. This is due to a mis-segregation (nondisjunction) during gametogenesis. The mother has a higher (10%) risk of transmission than the father (1%). Robertsonian translocations involving chromosome 14 also carry a slight risk of uniparental disomy 14 due to trisomy rescue.
Role in disease
Some human diseases caused by translocations are:
- Cancer: Several forms of cancer are caused by acquired translocations (as opposed to those present from conception); this has been described mainly in leukemia (acute myelogenous leukemia and chronic myelogenous leukemia). Translocations have also been described in solid malignancies such as Ewing's sarcoma.
- Infertility: One of the would-be parents carries a balanced translocation, where the parent is asymptomatic but conceived fetuses are not viable.
- Down syndrome is caused in a minority (5% or less) of cases by a Robertsonian translocation of the chromosome 21 long arm onto the long arm of chromosome 14.[7]
- Chromosomal translocations between the sex chromosomes can also result in a number of genetic conditions, such as XX male syndrome: caused by a translocation of the SRY gene from the Y to the X chromosome