
W2018_Bis2A_Lecture12_reading
- Page ID
- 10604
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)ATP
An important chemical compound is adenosine triphospate (ATP). The main cellular role of ATP is as a "short-term" energy transfer device for the cell. The hydrolysis reactions that liberate one or more of ATP's phosphates are exergonic and many, many cellular proteins have evolved to interact with ATP in ways that help facilitate the transfer of energy from hydrolysis to myriad other cellular functions. In this way, ATP is often called the “energy currency” of the cell: it has reasonably fixed values of energy to transfer to or from itself and can exchange that energy between many potential donors and acceptors. We will see many examples of ATP "at work" in the cell, so be looking for them. As you see them, try to think of them as functional examples of Nature's uses for ATP that you could be expected to see in another reaction or context.
ATP structure and function
At the heart of ATP is the nucleotide called adenosine monophosphate (AMP). Like the other nucleotides, AMP is composed of a nitrogenous base (an adenine molecule) bonded to a ribose molecule and a single phosphate group. The addition of a second phosphate group to this core molecule results in the formation of adenosine diphosphate (ADP); the addition of a third phosphate group forms adenosine triphosphate (ATP).

Figure 1. ATP (adenosine triphosphate) has three phosphate groups that can be removed by hydrolysis to form ADP (adenosine diphosphate) or AMP (adenosine monophosphate).
The phosphorylation (or condensation of phosphate groups onto AMP) is an endergonic process. By contrast, the hydrolysis of one or two phosphate groups from ATP, a process called dephosphorylation, is exergonic. Why? Let's recall that the terms endergonic and exergonic refer to the sign on the difference in free energy of a reaction between the products and reactants, ΔG. In this case we are explicitly assigning direction to the reaction, either in the direction of phosphorylation or dephosphorylation of the nucleotide. In the phosphorylation reaction the reactants are the nucleotide and an inorganic phosphate while the products are a phosphorylated nucleotide and WATER. In the dephosphorylation/hydrolysis reaction, the reactants are the phosphorylated nucleotide and WATER while the products are inorganic phosphate and the nucleotide minus one phosphate.
Since Gibbs free energy is a state function, it doesn't matter how the reaction happens; you just consider the beginning and ending states. As an example, let's examine the hydrolysis of ATP. The reactants ATP and water are characterized by their atomic makeup and the kinds of bonds between the constituent atoms. Some free energy can be associated with each of the bonds and their possible configurations—likewise for the products. If we examine the reaction from the standpoint of the products and reactants and ask "how can we recombine atoms and bonds in the reactants to get the products?," we find that a phosphoanhydride bond between an oxygen and a phosphorus must be broken in the ATP, a bond between an oxygen and hydrogen must be broken in the water, a bond must be made between the OH (that came from the splitting of water) and the phosphorus (from the freed PO3-2), and a bond must be formed between the H (derived from the splitting of water) and the terminal oxygen on the phosphorylated nucleotide. It is the sum of energies associated with all of those bond rearrangements (including those directly associated with water) that makes this reaction exergonic. A similar analysis could be made with the reverse reaction.
Possible Exercise
Use the figure of ATP above and your knowledge of what a water molecule looks like to draw a figure of the reaction steps described above: breaking of the phosphoanhydride bond, breaking of the water, and formation of new bonds to form ADP and inorganic phosphate. Track the atoms in different colors if that helps.
Is there something special about the specific bonds involved in these molecules? Much is made in various texts about the types of bonds between the phosphates of ATP. Certainly, the properties of the bonds in ATP help define the molecule's free energy and reactivity. However, while it is appropriate to apply concepts like charge density and availability of resonance structures to this discussion, trotting these terms out as an "explanation" without a thorough understanding of how these factors influence the free energy of the reactants is a special kind of hand-waving that we shouldn't engage in. Most BIS2A students have not had any college chemistry and those who have are not likely to have discussed those terms in any meaningful way. So, explaining the process using the ideas above only gives a false sense of understanding, assigns some mystical quality to ATP and its "special" bonds that don't exist, and distracts from the real point: the hydrolysis reaction is exergonic because of the properties of ATP and ALSO because of the chemical properties of water and those of the reaction products. For this class, it is sufficient to know that dedicated physical chemists are still studying the process of ATP hydrolysis in solution and in the context of proteins and that they are still trying to account for the key enthalpic and entropic components of the component free energies. We'll just need to accept a certain degree of mechanistic chemical ignorance and be content with a description of gross thermodynamic properties. The latter is perfectly sufficient to have deep discussions about the relevant biology.
"High-Energy" bonds
What about the term "high-energy bonds" that we so often hear associated with ATP? If there is nothing "special" about the bonds in ATP, why do we always hear the term "high-energy bonds" associated with the molecule? The answer is deceptively simple. In biology the term "high-energy bond" is used to describe an exergonic reaction involving the hydrolysis of the bond in question that results in a "large," negative change in free energy. Remember that this change in free energy does not only have to do with the bond in question but rather the sum of all bond rearrangements in the reaction. What constitutes a large change? It is a rather arbitrary assignment usually associated with an amount of energy associated with the types of anabolic reactions we typically observe in biology. If there is something special about the bonds in ATP, it is not uniquely tied to the free energy of hydrolysis, as there are plenty of other bonds whose hydrolysis results in greater negative differences in free energy.
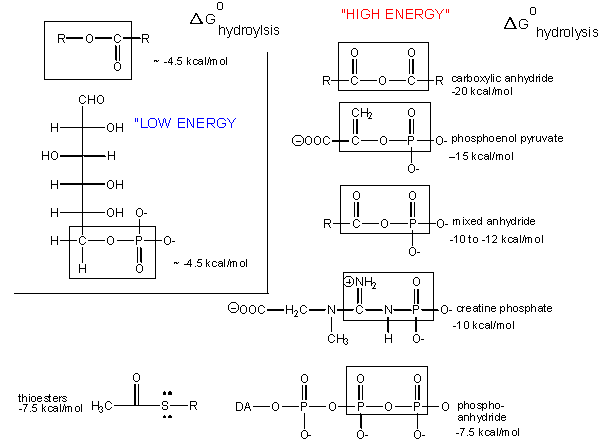
Figure 2. The free energy of hydrolysis of different types of bonds can be compared to that of the hydrolysis of ATP. Source: http://bio.libretexts.org/Core/Biochemistry/Oxidation_and_Phosphorylation/ATP_and_Oxidative_Phosphorylation/Properties_of_ATP
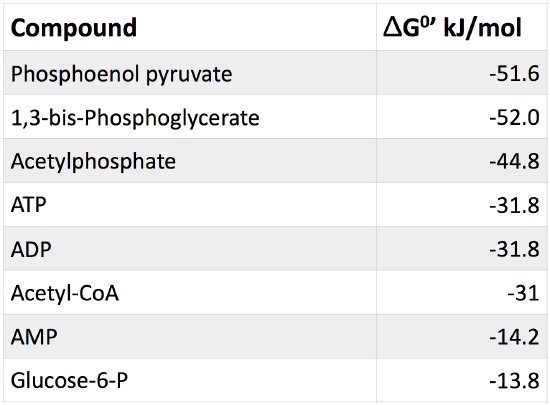
Table 1. Table of common cellular phosphorylated molecules and their respective free energies of hydrolysis.
External link discussing the energetics of coupling ATP hydrolysis to other reactions
The cycling of ATP pools
Estimates for the number of ATP molecules in a typical human cell range from ~3x107 (~5x10-17 moles ATP/cell) in a white blood cell to 5x109 (~9x10-15 moles ATP/cell) in an active cancer cell. While these numbers might seem large, and already amazing, consider that it is estimated that this pool of ATP turns over (becomes ADP and then back to ATP) 1.5 x per minute. Extending this analysis yields the estimate that this daily turnover amounts to roughly the equivalent of one body weight of ATP getting turned over per day. That is, if no turnover/recycling of ATP happened, it would take one body weight worth of ATP for the human body to function, hence our previous characterization of ATP as a "short-term" energy transfer device for the cell.
While the pool of ATP/ADP may be recycled, some of the energy that is transferred in the many conversions between ATP, ADP, and other biomolecules is also transferred to the environment. In order to maintain cellular energy pools, energy must transfer in from the environment as well. Where does this energy come from? The answer depends a lot on where energy is available and what mechanisms Nature has evolved to transfer energy from the environment to molecular carriers like ATP. In nearly all cases, however, the mechanism of transfer has evolved to include some form of redox chemistry.
In this and the sections that follow we are concerned with learning some critical examples of energy transfer from the environment, key types of chemistry and biological reactions involved in this process, and key biological reactions and cellular components associated with energy flow between different parts of the living system. We focus first on reactions involved in the (re)generation of ATP in the cell (not those involved in the creation of the nucleotide per se but rather those associated with the transfer of phosphates onto AMP and ADP).
Video link
For another perspective - including places you'll see ATP in Bis2a, take a look at this video (10 minutes) by clicking here.

How do cells generate ATP?
A variety of mechanisms have emerged over the 3.25 billion years of evolution to create ATP from ADP and AMP. The majority of these mechanisms are modifications on two themes: direct synthesis of ATP or indirect synthesis of ATP with two basic mechanisms known respectively as substrate level phosphorylation (SLP) and oxidative phosphorylation. Both mechanisms rely on biochemical reactions that transfer energy from some energy source to ADP or AMP to synthesize ATP. These topics are substantive, so they will be discussed in detail in the next few modules.
Introduction to Mobile Energy Carriers
Section Summary
Energy is moved around and transferred within the cell in a variety of ways. One critical mechanism that nature has developed is the use of recyclable molecular energy carriers. While there are several major recyclable energy carriers, they all share some common functional features:
Properties of Key Cellular Molecular Energy Carriers
- We think of the energy carriers as existing in "pools" of available carriers. One could, by analogy, consider these mobile energy carriers analogous to the delivery vehicles of parcel carriers—the company has a certain "pool" of available vehicles at any one time to pickup and make deliveries.
- Each individual carrier in the pool can exist in one of multiple distinct states: it is either carrying a "load" of energy, a fractional load, or is "empty". The molecule can interconvert between "loaded" and empty and thus can be recycled. Again by analogy, the delivery vehicles can be either carrying packages or be empty and switch between these states.
- The balance or ratio in the pool between "loaded" and "unloaded" carriers is important for cellular function, is regulated by the cell, and can often tell us something about the state of a cell. Likewise, a parcel carrier service keeps close tabs on how full or empty their delivery vehicles are—if they are too full, there may be insufficient "empty" trucks to pick up new packages; if they are too empty, business must not be going well or it is shut down. There is an appropriate balance for different situations.
In this course, we will examine two major types of molecular recyclable energy carriers: (1) the adenine nucleotides, specifically: nicotinamide adenine dinucleotide (NAD+), a close relative, nicotinamide adenine dinucleotide phosphate (NADP+), and flavin adenine dinucleotide (FAD2+) and (2) nucleotide mono-, di-, and triphosphates, with particular attention paid to adenosine triphosphate (ATP). Each of these two types of molecules is involved in energy transfer that involves different classes of chemical reactions. Adenine nucleotides are primarily associated with redox chemistry, while the nucleotide triphosphates are associated with transfers of energy that are linked to the hydrolysis or condensation of inorganic phosphates.
Red/ox chemistry and electron carriers
The oxidation of, or removal of an electron from, a molecule (whether accompanied with the removal of an accompanying proton or not) results in a change of free energy for that molecule—matter, internal energy, and entropy have all changed in the process. Likewise, the reduction of (the gain of electron on) a molecule also changes its free energy. The magnitude of change in free energy and its direction (positive or negative) for a red/ox reaction dictates the spontaneity of the reaction and how much energy is transferred. In biological systems, where a great deal of energy transfer happens via red/ox reactions, it is important to understand how these reactions are mediated and to begin to start considering ideas or hypotheses for why these reactions are mediated in many cases by a small family of electron carriers.
Note: possible discussion
Relate the burning of (the full oxidation of the sugar in) a gummy bear with the last paragraph above. What does that demonstration have to do with our upcoming discussion on red/ox carriers? There is some mention above already—can you find it?

Note: possible discussion
The problem alluded to in the previous discussion question is a great place to start bringing in the design challenge rubric. If you recall, the first step of the rubric asks that you define a problem or question. In this case, let's imagine that there is a problem to define for which the mobile electron carriers below helped Nature solve.
***Remember, evolution DOES NOT forward-engineer solutions to problems, but in retrospect, we can use our imagination and logic to infer that what we see preserved by natural selection provided a selective advantage, because the natural innovation "solved" a problem that limited success.***
Design challenge for red/ox carriers
- What was a problem(s) that the evolution of mobile electron/red/ox carriers helped solve?
- The next step of the design challenge asks you to identify criteria for successful solutions. What are criteria for success in the problem you've identified?
- Step 3 in the design challenge asks you to identify possible solutions. Well, here Nature has identified some for us—we consider three in the reading below. It looks like Nature is happy to have multiple solutions to the problem.
- The penultimate step of the design challenge rubric asks you to evaluate the proposed solutions against the criteria for success. This should make you think/discuss about why there are multiple different electron carriers. Are there different criteria for success? Are they each solving slightly different problems? What do you think? Be on the lookout as we go through metabolism for clues.
NAD+/H and FADH/H2
In living systems, a small class of compounds function as electron shuttles: they bind and carry electrons between compounds in different metabolic pathways. The principal electron carriers we will consider are derived from the B vitamin group and are derivatives of nucleotides. These compounds can be both reduced (that is, they accept electrons) or oxidized (they lose electrons) depending on the reduction potential of a potential electron donor or acceptor that they might transfer electrons to and from. Nicotinamide adenine dinucleotide (NAD+) (the structure is shown below) is derived from vitamin B3, niacin. NAD+ is the oxidized form of the molecule; NADH is the reduced form of the molecule after it has accepted two electrons and a proton (which together are the equivalent of a hydrogen atom with an extra electron).
We are expecting you to memorize the two forms of NAD+/NADH, know which form is oxidized and which is reduced, and be able to recognize either form on the spot in the context of a chemical reaction.
NAD+ can accept electrons from an organic molecule according to the general equation:

Here is some vocabulary review: when electrons are added to a compound, the compound is said to have been reduced. A compound that reduces (donates electrons to) another is called a reducing agent. In the above equation, RH is a reducing agent, and NAD+ is reduced to NADH. When electrons are removed from a compound, it becomes oxidized. A compound that oxidizes another is called an oxidizing agent. In the above equation, NAD+ is an oxidizing agent, and RH is oxidized to R.
You need to get this down! We will (a) test specifically on your ability to do so (as "easy" questions), and (b) we will use the terms with the expectation that you know what they mean and can relate them to biochemical reactions correctly (in class and on tests).
You will also encounter a second variation of NAD+, NADP+. It is structurally very similar to NAD+, but it contains an extra phosphate group and plays an important role in anabolic reactions, such as photosynthesis. Another nucleotide-based electron carrier that you will also encounter in this course and beyond, flavin adenine dinucleotide (FAD+), is derived from vitamin B2, also called riboflavin. Its reduced form is FADH2. Learn to recognize these molecules as electron carriers as well.

Figure 1. The oxidized form of the electron carrier (NAD+) is shown on the left, and the reduced form (NADH) is shown on the right. The nitrogenous base in NADH has one more hydrogen ion and two more electrons than in NAD+.
NAD+ is used by the cell to "pull" electrons off of compounds and to "carry" them to other locations within the cell; thus it is called an electron carrier. NAD+/H compounds are used in many of the metabolic processes we will discuss in this class. For example, in its oxidized form, NAD+ is used as a reactant in glycolysis and the TCA cycle, whereas in its reduced form (NADH), it is a reactant in fermentation reactions and the electron transport chain (ETC). Each of these processes will be discussed in later modules.
Energy story for a red/ox reaction
***As a rule of thumb, when we see NAD+/H as a reactant or product, we know we are looking at a red/ox reaction.***
When NADH is a product and NAD+ is a reactant, we know that NAD+ has become reduced (forming NADH); therefore, the other reactant must have been the electron donor and become oxidized. The reverse is also true. If NADH has become NAD+, then the other reactant must have gained the electron from NADH and become reduced.
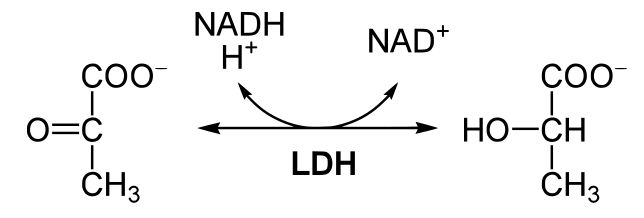
Figure 2. This reaction shows the conversion of pyruvate to lactic acid coupled with the conversion of NADH to NAD+. Source: https://en.wikibooks.org/wiki/Structural_Biochemistry/Enzyme/sequential_reactions
In the figure above, we see pyruvate becoming lactic acid, coupled with the conversion of NADH into NAD+. This reaction is catalyzed by LDH. Using our "rule of thumb" above, we categorize this reaction as a red/ox reaction. NADH is the reduced form of the electron carrier, and NADH is converted into NAD+. This half of the reaction results in the oxidation of the electron carrier. Pyruvate is converted into lactic acid in this reaction. Both of these sugars are negatively charged, so it would be difficult to see which compound is more reduced using the charges of the compounds. However, we know that pyruvate has become reduced to form lactic acid, because this conversion is coupled to the oxidation of NADH into NAD+. But how can we tell that lactic acid is more reduced than pyruvate? The answer is to look at the carbon-hydrogen bonds in both compounds. As electrons are transferred, they are often accompanied by a hydrogen atom. There is a total of three C-H bonds in pyruvate, and there is a total of four C-H bonds in lactic acid. When we compare these two compounds in the before and after states, we see that lactic acid has one more C-H bond; therefore, lactic acid is more reduced than pyruvate. This holds true for multiple compounds. For example, in the figure below, you should be able to rank the compounds from most to least reduced using the C-H bonds as your guide.

Figure 3. Above are a series of compounds than can be ranked or reorganized from most to least reduced. Compare the number of C-H bonds in each compound. Carbon dioxide has no C-H bonds and is the most oxidized form of carbon we will discuss in this class. Answer: the most reduced is methane (compound 3), then methanol (4), formaldehyde (1), carboxylic acid (2), and finally carbon dioxide (5).

Figure 4. This reaction shows the conversion of G3P, NAD+, and Pi into NADH and 1,3-BPG. This reaction is catalyzed by glyceraldehyde-3-phosphate dehydrogenase.
Energy story for the reaction catalyzed by glyceraldehyde-3-phosphate dehydrogenase:
Lets make an energy story for the reaction above.
First, lets characterize the reactants and products. The reactants are glyceraldehyde-3-phosphate (a carbon compound), Pi (inorganic phosphate), and NAD+. These three reactants enter into a chemical reaction to produce two products, NADH and 1,3-bisphosphoglycerate. If you look closely, you can see that the 1,3-BPG contains two phosphates. This is important when we are double checking that no mass has been lost. There are two phosphates in the reactants, so there must be two phosphates in the products (conservation of mass!). You can double check that all the other atoms are also accounted for. The enzyme that catalyzes this reaction is called glyceraldehyde-3-phosphate dehydrogenase. The standard free energy change of this reaction is ~6.3 kJ/mol, so under standard conditions, we can say that the free energy of the products is higher than that of the reactants and that this reaction is not spontaneous under standard conditions.
What can we say about this reaction when it is catalyzed by glyceraldehyde-3-phosphate dehydrogenase?
This is a red/ox reaction. We know that because we have produced a reduced electron carrier (NADH) as a product and NAD+ is a reactant. Where did the electron come from to make NADH? The electron must have come from the other reactant (the carbon compound).
Note: recommended discussion
We will spend some time examining the reaction catalyzed by glyceraldehyde-3-phosphate dehydrogenase in more detail as we move through the lectures and text. The first thing to discuss here is that the figure above is a highly simplified or condensed version of the steps that take place—one could in fact break that reaction above into TWO conceptual reactions. Can you imagine what those two "subreactions" might be? Discuss amongst yourselves.
Note: recommended discussion
The text above notes that the standard change in free energy for this complex reaction is ~+6.3 kJ/mol. Under standard conditions, this reaction is NOT spontaneous. However, this is one of the key reactions in the oxidation of glucose. It needs to GO in the cell. The questions are as follows: why is it important to note things like "standard change of free energy" or "under standard conditions" when reporting that ΔG°? What could possibly be going on in the cell to make what is under standard conditions an endergonic reaction "go"?