
18: Transcriptional regulation after initiation
- Page ID
- 389

Although regulation of the initiation of transcription appears to be a dominant factor in control of expression of many genes, the importance of regulation after initiation is becoming better appreciated in an increasing number and variety of systems. The classic systems in which these issues have been explored are antitermination in bacteriophage l and in attenuation of transcription in bacterial biosynthetic operons, in particular the trp operon in E. coli. Although some of the mechanistic details may be peculiar to bacteria, especially the need for coupled transcription and translation in the trpattenuation system, the phenomenon of regulation after initiation is seen in a wide variety of organisms, ranging from bacteria to humans. Some of this work was discussed in the sections on elongation of transcription in Chapter II of Part Three.
Introduction
Both systems discussed in this chapter control the frequency of termination of transcription. Antitermination in bacteriophage l can prevent RNA polymerase from stopping at r-dependent terminators, thus leading to transcription of downstream genes. Attenuation in the trp operon also controls the frequency at which RNA polymerase stops at an early terminator in the operon, hence regulating the transcription of downstream genes. In contrast to the system in l, attenuation in trp regulates termination at a r-independentterminator.
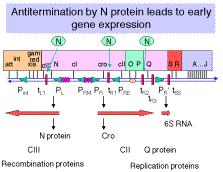
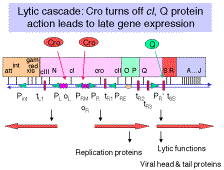
Antitermination in Bacteriophage l
Just to quickly review one of the points in Chapter III, antitermination occurs at two different times in the llife cycle. The N protein allows read-through transcription in the shift from immediate-early to early transcription, and the Q protein allows read-through transcription of the late genes.
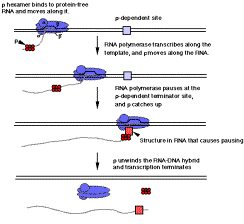
Recall from Part Three of the text that r-dependent terminators do not have a well-conserved sequence or secondary structure. Also, the protein r tracks along protein-free regions of the RNA until it hits a paused transcriptional complex at a r-dependent site, at which point its RNA helicase activity can cause termination and dissociation of the polymerase and transcript from the template DNA.

Sites on the DNA needed for antitermination in bacteriophage l. nutsites (N utilization sites) for pN, qutsite for pQ and the nutsites are within the transcription unit, not at the promoter and not at the terminator.
- nutLis in the 5' untranslated region of the Ngene, and nutRis in the 3' untranslated region of the crogene.
- In both cases, the nutsite precedes the terminator at which pN will act.

Both nutLand nutR are 17 bp sequences with a dyad symmetry.
5' AGCCCTGAARAAGGGCA
TCGGGACTTYTTCCCGT 5'
The protein pN recognizes the nutsite and binds to RNA polymerase as it transcribes through the site. The complex of pN with the RNA polymerases is highly processive and overrides the efforts of r at the terminator.
E. coli (host) proteins needed for action of pN. These were isolated as host functions that when mutated prevented action of pN. NusA (encoded by nusA, for N utilization substance, complementation group A) is the best characterized.
- Can form part of the transcription complex
- Has been proposed to bind to the core RNA polymerase after s dissociates.
- Can also bind pN.
- Model:
NusA binds the core polymerase after s dissociates. As this complex transcribes through a nutsite, pN binds also. The complex a2bb'-NusA-pN prevents r-dependent termination at tR1, tR2, and tL1.
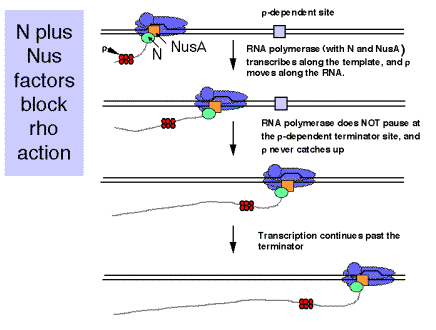
Several other nusgenes have been identified. NusG is the bacterial homolog of a family of conserved proteins involved in elongation. It is homologous to the large subunit of DSIF, which is an elongation factor in mammals. DSIF is the DRB-sensitivity inducing factor. Current studies implicate it in both negative and positive effects on elongation. It has two subunits, one of 160 kDa that is homologous to the yeast transcriptional regulatory protein Spt5, and one of 14 kDa that is homologous to the yeast Spt4 protein. Another nusgene encodes a ribosomal protein. Much more needs to be learned about both termination and antitermination. The nusphenotype of mutations in a gene encoding a ribosomal protein suggests that translation is also coupled to this process.
Components of the E. coli trp operon
The trpoperon encodes the enzymes required for biosynthesis of tryptophan. More specifically, its five genes (trpEDCBA) encode five subunits of proteins that in total catalyze five enzymatic steps, converting chorismic acid to tryptophan. However, there is not a 1:1 correspondence between a cistron and an enzyme. For example, trpBand trpAencode, respectively, the b and a subunits of tryptophan synthase, which catalyzes the replacement of glycerol-3-phosphate from indole-3-glycerol-phosphate with serine to form tryptophan, with glyceraldehyde-3-phosphate as the other product
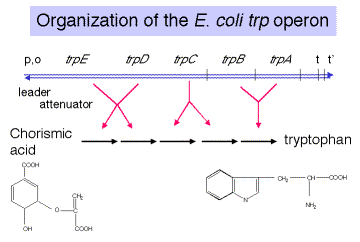
A leader sequence separates the promoter and operator from the first structural gene of the operon, trpE. An attenuator of transcription follows the leader. As we will see in more detail below, the efficiency of "premature" termination at this attenuator is determined by the extent of translation of the leader, which in turn is determined by the availability of Trp-tRNAtrp. This is an important part of the regulation of the operon. Two terminators of transcription follow the structural genes, one dependent on r and one independent of r.
Modes of regulation: turn operon off in presence of Trp
Repressor-operator: requires a protein binding to a specific site in the presence of Trp to decrease the efficiency of initiation of transcription. Attenuation: the elongation (and termination) of transcription by RNA polymerase is linked to the progress of translation by a ribosome. In the presence of Trp, the translation by the ribosome causes transcription of the subsequent genes in the operon to terminate.
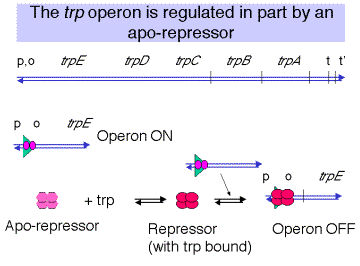
Repressor: apo-repressor and co-repressor (Trp)
The apo-repressor is encoded by trpRat a distant locus. The apo-repressor is a homo-tetramer. It has a high affinity for the operator only when it is bound by the amino acid Trp, which serves as a co-repressor. Thus the active repressor is a tetramer of (formerly apo-) repressor in complex with Trp. The active repressor binds to the operator to prevent initiation of transcription. The operator overlaps the promoter, including the -10 region of the promoter. It has a dyad axis of symmetry.
Attenuation
The attenuator is a conditional transcriptional terminator used to regulate expression of biosynthetic operons in bacteria. It is upstream of the structural genes trpEDCBA and is a r-independent termination site. Its ability to terminate transcription is dependent on its ability to form the stem of duplex RNA that is characteristic of r-independent termination sites.
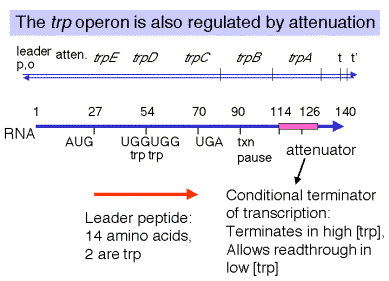
The fraction of transcripts that read through the attenuator is determined by the [Trp-tRNAtrp]. The concentration of charged tRNAs is a measure of the amount of Trp available for protein synthesis. If most tRNAtrp is charged, there is an abundance of Trp, and the cell does not need to make more. Low [Trp-tRNAtrp] allows read-through transcription through the attenuator, so that trpEDCBA is expressed and high [Trp-tRNAtrp] causes termination of transcription at the attenuator.
The [Trp-tRNAtrp] determines the progress of ribosomes as they translate a short leader peptide. The leader peptide is a short 14 amino acid polypeptide encoded by trpL. Two codons for Trp are in the leader, and the progress of ribosomes past these Trp codons will be determined by the availability of Trp-tRNAtrp. When the concentration of tryptophanyl-tRNA is high, translation of the trpleader will be completed, but when it is low, translation will stall at the tryptophan codons.
The extent of progress of the ribosomes determines the secondary structures formed in the leader RNA. When the [Trp-tRNAtrp] is high, the ribosomes translate past the Trp codons to complete the synthesis leader of the peptide. This allows the nascent RNA to form the structure for r-independent terminator. Thus transcription terminates before the RNA polymerase reaches trpEDCBA. When the [Trp-tRNAtrp] is low, the ribosomes stall at the Trp codons, which prevents formation of the secondary structures in the RNA necessary for termination at the attenuator. Thus read-through transcription continues through trpEDCBAand the operon is expressed, so that more Trp is made.
| [trp-tRNA] | translation of trpL | secondary structures formed in RNA | Attenuator | Operon |
|---|---|---|---|---|
| High | complete | 3-4 stem | terminate transcription | OFF |
| Low | stalls at Trp codons | 2-3 stem | allow read-through transcription | ON |
Alternative base-paired structures in leader RNA. Four regions of the leader RNA can be involved in secondary structure formation, in particular base-paired stems; these are referred to simply as regions 1, 2, 3, and 4. Potentially, 1 can pair with 2, 2 can pair with 3, and 3 can pair with 4.
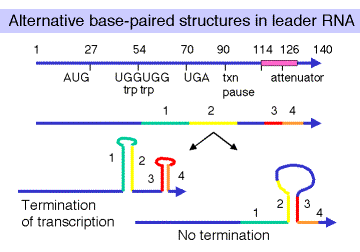
A stem formed by pairing between 3 and 4 makes a G+C rich stem followed by U's, which is sufficient for r-independent termination of transcription. When the [Trp-tRNAtrp] is high, the 3-4 base-paired structure forms, and transcription terminates at the attenuator. This turns the operon OFF. The formation of a base-paired stem between regions 2 and 3 precludes formation of the 3-4 terminator, and transcription will continue into the structural genes trpEDCBA. This turns the operon ON.
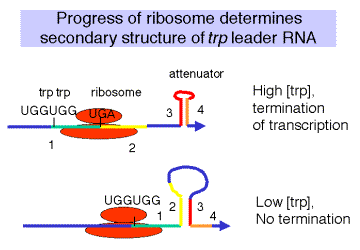
The choice between a 2-3 stem or a 3-4 stem is dictated by the progress of the ribosome. If the ribosome can translate past the Trp codons (when the [Trp-tRNAtrp] is high), then it will reach a natural translation termination codon. When the ribosome is in that position, region 2 of the leader RNA is covered by the ribosome, so the 2-3 stem cannot form but the 3-4 stem can. This generates the secondary structure needed for termination of transcription at the attenuator. In contrast, if the ribosome stalls at the Trp codons in the leader, because the [Trp-tRNAtrp] is low, then region 2 of the leader RNA is not covered by the ribosome. It can then base pair with region 3. This prevents formation of the 3-4 terminator, and RNA polymerase can continue elongation through trpEDCBA.
Mutational Analysis (selected examples)
- Translation of trpLis needed for regulation by attenuation. Mutation of the AUG for initiation of translation of the leader RNA prevents transcription past the attenuator. In the absence of translation, both the 1-2 and 3-4 stems can form. The latter 3-4 stem is the terminator.
- Charged tRNAtrp is required for regulation. Mutation of the genes for tRNAtrp or Trp-tRNAtrp synthetase leads to constitutive expression of trpEDCBA. In these mutants, translation will stall at Trp codons regardless of the intracellular [Trp], and no terminator will form at the attenuator.
- Specific secondary structures in the trpleader RNA are needed for regulation. E.g. mutations that decrease the number of base pairs between the 3 and 4 regions will decrease the amount of transcriptional termination (i.e. increase expression of the operon). Compensatory mutations that increase the number of base pairs between 3 and 4 will suppress the original mutations.
Attenuation requires coupled transcription and translation
Requires no regulatory proteins: charging of cognate tRNA is the regulatory signal. Need a transcriptional pause site at +90 to allow the ribosomes to catch up with the RNA polymerase and thereby affect the secondary structures in the nascent RNA.
Attenuation is a common mechanism for regulating biosynthetic operons
Many operons that encode the enzymes catalyzing biosynthesis of amino acids are regulated by attenuation. In each case, the leader polypeptide is rich in the amino acid that is the product of the pathway, e.g. his, phe, leu, thr, ilv.
Additional readings
- Friedman, D.I. and Count, D.L. (1995) Transcriptional antitermination: The lambda paradigm updated. Molecular Microbiology 18: 191-200.
- Henkin, T. (2000) Transcriptional termination in bacteria. Current Opinions in Microbiology 3: 149-153.
- Gusarov, I. and Nudler, E. (2001) Control of intrinsic transcriptional termination by N and NusA: The basic mechanism. Cell 107: 437-449.