
2.5: B-Form, A-Form, and Z-Form of DNA
- Page ID
- 307

Three major forms of DNA are double stranded and connected by interactions between complementary base pairs. These are terms A-form, B-form,and Z-form DNA.
B-form DNA
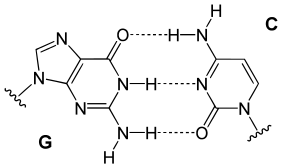
The information from the base composition of DNA, the knowledge of dinucleotide structure, and the insight that the X‑ray crystallography suggested a helical periodicity were combined by Watson and Crick in 1953 in their proposed model for a double helical structure for DNA. They proposed two strands of DNA -- each in a right‑hand helix -- wound around the same axis. The two strands are held together by H‑bonding between the bases (in anti conformation) as shown in Figure \(\PageIndex{1}\).
Major groove Major groove

Minor groove Minor groove
Figure \(\PageIndex{1}\): (left) An A:T base pair and (right) a G:C base pair
Bases fit in the double helical model if pyrimidine on one strand is always paired with purine on the other. From Chargaff's rules, the two strands will pair A with T and G with C. This pairs a keto base with an amino base, a purine with a pyrimidine. Two H‑bonds can form between A and T, and three can form between G and C. This third H-bond in the G:C base pair is between the additional exocyclic amino group on G and the C2 keto group on C. The pyrimidine C2 keto group is not involved in hydrogen bonding in the A:T base pair.
These are the complementary base pairs. The base‑pairing scheme immediately suggests a way to replicate and copy the the genetic information.
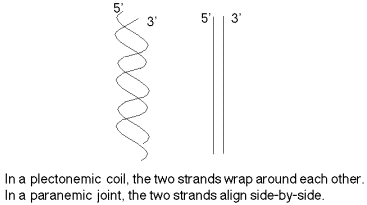
The two strands are not in a simple side‑by‑side arrangement, which would be called a paranemic joint (Figure \(\PageIndex{3}\)). (This will be encountered during recombination in Chapter 8.) Rather the two strands are coiled around the same helical axis and are intertwined with themselves (which is referred to as a plectonemic coil). One consequence of this intertwining is that the two strands cannot be separated without the DNA rotating, one turn of the DNA for every "untwisting" of the two strands.
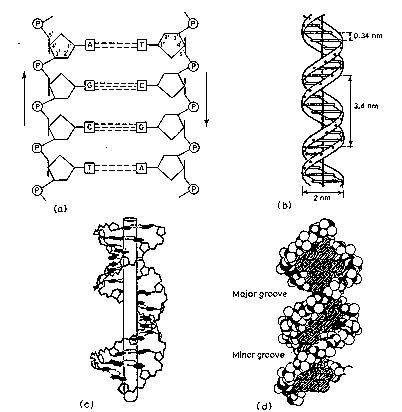
Dimensions of B-form (the most common) of DNA
- 0.34 nm between bp, 3.4 nm per turn, about 10 bp per turn
- 1.9 nm (about 2.0 nm or 20 Angstroms) in diameter
Major and minor groove
The major groove is wider than the minor groove in DNA (Figure \(\PageIndex{2d}\)), and many sequence specific proteins interact in the major groove. The N7 and C6 groups of purines and the C4 and C5 groups of pyrimidines face into the major groove, thus they can make specific contacts with amino acids in DNA-binding proteins. Thus specific amino acids serve as H‑bond donors and acceptors to form H-bonds with specific nucleotides in the DNA. H‑bond donors and acceptors are also in the minor groove, and indeed some proteins bind specifically in the minor groove. Base pairs stack, with some rotation between them.
A‑form nucleic acids and Z‑DNA
Three different forms of duplex nucleic acid have been described. The most common form, present in most DNA at neutral pH and physiological salt concentrations, is B-form. That is the classic, right-handed double helical structure we have been discussing. A thicker right-handed duplex with a shorter distance between the base pairs has been described for RNA-DNA duplexes and RNA-RNA duplexes. This is called A-form nucleic acid.
A third form of duplex DNA has a strikingly different, left-handed helical structure. This Z DNA is formed by stretches of alternating purines and pyrimidines, e.g. GCGCGC, especially in negatively supercoiled DNA. A small amount of the DNA in a cell exists in the Z form. It has been tantalizing to propose that this different structure is involved in some way in regulation of some cellular function, such as transcription or regulation, but conclusive evidence for or against this proposal is not available yet.
Differences between A-form and B-form nucleic acid
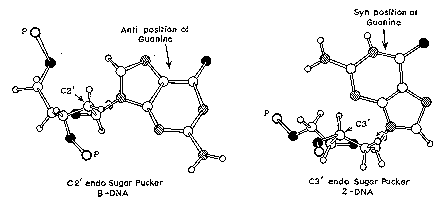
The major difference between A-form and B-form nucleic acid is in the conformation of the deoxyribose sugar ring. It is in the C2' endoconformation for B-form, whereas it is in the C3' endoconformation in A-form. As shown in Figure \(\PageIndex{4}\), if you consider the plane defined by the C4'-O-C1' atoms of the deoxyribose, in the C2' endoconformation, the C2' atom is above the plane, whereas the C3' atom is above the plane in the C3' endoconformation. The latter conformation brings the 5' and 3' hydroxyls (both esterified to the phosphates linking to the next nucleotides) closer together than is seen in the C2' endoconfromation (Figure 2.16). Thus the distance between adjacent nucleotides is reduced by about 1 Angstrom in A-form relative to B-form nucleic acid (Figure \(\PageIndex{4}\)).
A second major difference between A-form and B-form nucleic acid is the placement of base-pairs within the duplex. In B-form, the base-pairs are almost centered over the helical axis (Figure \(\PageIndex{4}\)), but in A-form, they are displaced away from the central axis and closer to the major groove. The result is a ribbon-like helix with a more open cylindrical core in A-form.
Z-form DNA
Z-DNA is a radically different duplex structure, with the two strands coiling in left-handed helices and a pronounced zig-zag (hence the name) pattern in the phosphodiester backbone. As previously mentioned, Z-DNA can form when the DNA is in an alternating purine-pyrimidine sequence such as GCGCGC, and indeed the G and C nucleotides are in different conformations, leading to the zig-zag pattern. The big difference is at the G nucleotide. It has the sugar in the C3' endoconformation (like A-form nucleic acid, and in contrast to B-form DNA) and the guanine base is in the synconformation. This places the guanine back over the sugar ring, in contrast to the usual anticonformation seen in A- and B-form nucleic acid. Note that having the base in the anticonformation places it in the position where it can readily form H-bonds with the complementary base on the opposite strand. The duplex in Z-DNA has to accomodate the distortion of this G nucleotide in the synconformation. The cytosine in the adjacent nucleotide of Z-DNA is in the "normal" C2' endo, anticonformation.
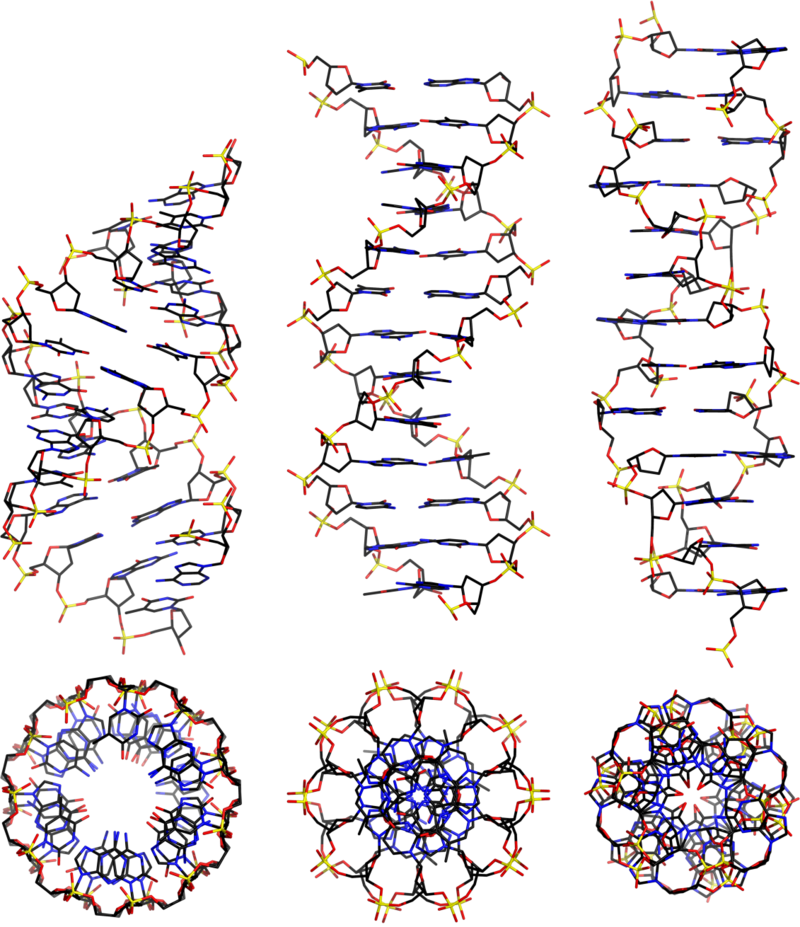
Even classic B-DNA is not completely uniform in its structure. X-ray diffraction analysis of crystals of duplex oligonucleotides shows that a given sequence will adopt a distinctive structure. These variations in B-DNA may differ in the propeller twist (between bases within a pair) to optimize base stacking, or in the 3 ways that 2 successive base pairs can move relative to each other: twist, roll, or slide.
| B-Form | A-Form | Z-Form | |
|---|---|---|---|
| helix sense | Right Handed | Right Handed | Left Handed |
| base pairs per turn | 10 | 11 | 12 |
| vertical rise per bp | 3.4 Å | 2.56 Å | 19 Å |
| rotation per bp | +36° | +33° | -30° |
| helical diameter | 19 Å | 19 Å | 19 Å |