
11.3E: Phagocytosis
- Page ID
- 3280

Briefly describe the role of the following as they relate to phagocytosis: - inflammation
- lymph nodules
- lymph nodes
- spleen
Describe the following steps in phagocytosis: - activation
- chemotaxis
- attachment (both unenhanced and enhanced)
- ingestion
- destruction
State what happens when either phagocytes are overwhelmed with microbes or they adhere to cells to large to be phagocytosed. Describe what causes most of the tissue destruction seen during microbial infections. Compare the oxygen-dependent and oxygen-independent killing systems of neutrophils and macrophages. Briefly describe the role of autophagy in removing intracellular microbes.
- activation
- chemotaxis
- attachment (both unenhanced and enhanced)
- ingestion
- destruction
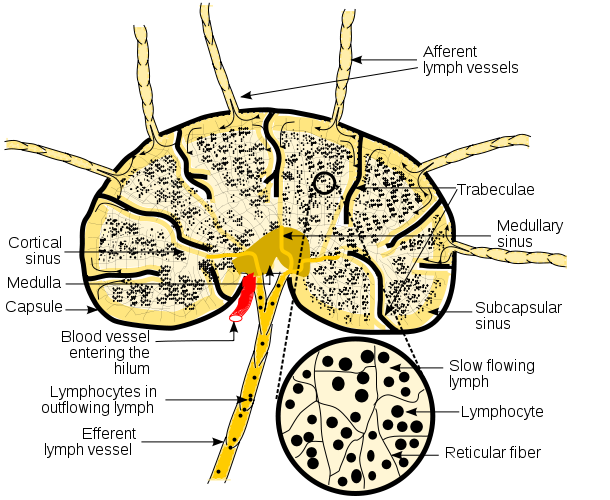
In addition, Langerhans' cells (immature dendritic cells) are located throughout the epithelium of the skin, the respiratory tract, and the gastrointestinal tract where in their immature form they are attached by long cytoplasmic processes. Upon capturing antigens through pinocytosis and phagocytosis and becoming activated by proinflammatory cytokines, the dendritic cells detach from the epithelium, enter lymph vessels, and are carried to regional lymph nodes. By the time they enter the lymph nodes, they have matured and are now able to present antigen to the ever changing populations of naive T-lymphocytes located in the cortex of the lymph nodes.
The spleen contains many reticular fibers that support fixed macrophages and dendritic cells, as well as ever changing populations of circulating B-lymphocytes and T-lymphocytes. Blood carries microorganisms to the spleen where they are filtered out and phagocytosed by the fixed macrophages and dendritic cells and presented to the circulating B-lymphocytes and T-lymphocytes to initiate adaptive immune responses. There are also specialized macrophages and dendritic cells located in the brain (microglia), lungs (alveolar macrophages), liver (Kupffer cells), kidneys (mesangial cells), bones (osteoclasts), and the gastrointestinal tract (peritoneal macrophages).
The Steps Involved in Phagocytosis
There are a number of distinct steps involved in phagocytosis:
Step 1: Activation of the Phagocyte
Resting phagocytes are activated by inflammatory mediators such as bacterial products (bacterial proteins, capsules, LPS, peptidoglycan, teichoic acids, etc.), complement proteins, inflammatory cytokines, and prostaglandins. As a result, the circulating phagocytes produce surface glycoprotein receptors that increase their ability to adhere to the inner surface of capillary walls, enabling them to squeeze out of the capillary and be attracted to the site of infection.
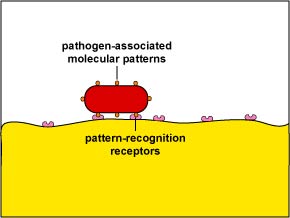
In addition, they produce endocytic pattern-recognition receptors that recognize and bind to pathogen-associated molecular patterns or PAMPs - components of common microbial molecules such as peptidoglycan, teichoic acids, lipopolysaccharide, and mannose-rich glycans that are not found in human cells - to attach the microbe to the phagocyte for what is called unenhanced attachment (discussed below). They also exhibit increased metabolic and microbicidal activity by increasing their production of ATPs, lysosomal enzymes, lethal oxidants, etc.
Step 2: Chemotaxis of Phagocytes (for wandering macrophages, neutrophils, and eosinophils)
Chemotaxis is the movement of phagocytes toward an increasing concentration of some attractant such as bacterial factors (bacterial proteins, capsules, LPS, peptidoglycan, teichoic acids, etc.), complement proteins (C5a), chemokines (chemotactic cytokines such as interleukin-8 secreted by various cells), fibrin split products, kinins, and phospholipids released by injured host cells.
| Movie showing chemotaxis by neutrophil. Chemotaxis of Neutrophils. © From Intimate Strangers: Unseen Life on Earth. Created by Mondo Media. Peter Baker, Executive Producer. Licensed for use, ASM MicrobeLibrary. |
| You Tube animation summarizing phagocytosis by a macrophage. |
| You Tube movie illustrating chemotaxis. |
Some microbes, such as the influenza A viruses, Mycobacterium tuberculosis, blood invasive strains of Neisseria gonorrhoeae, and Bordetella pertussis have been shown to block chemotaxis.
Step 3: Attachment of the Phagocyte to the Microbe or Cell
Attachment of microorganisms is necessary for ingestion. Attachment may be unenhanced or enhanced.
c. Extracellular trapping with NETs: In response to certain pathogen associated molecular patterns such as LPS, and certain cytokines such as IL-8, neutrophils release DNA and antimicrobial granular proteins. These neutrophil extracellular traps (NETs) bind to bacteria, prevent them from spreading, and kill them with antimicrobial proteins (see Figure \(\PageIndex{15}\) and Figure \(\PageIndex{16}\)).
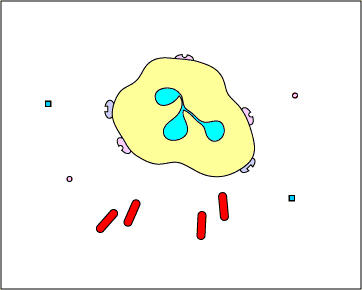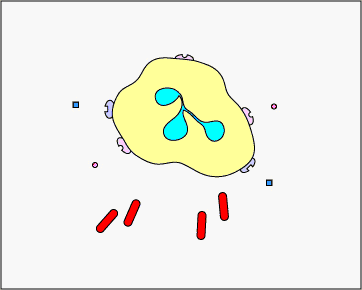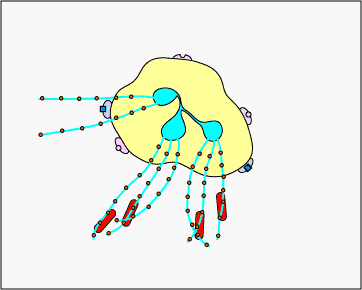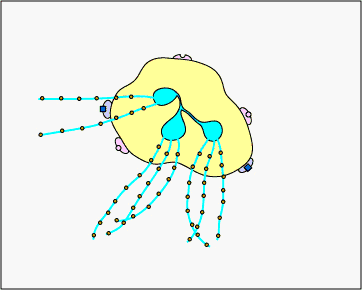
Neutrophil NETS Trapping and Killing Bacteria. In response to certain pathogen associated molecular patterns such as LPS, and certain cytokines such as IL-8, neutrophils release DNA and antimicrobial granular proteins. These neutrophil extracellular traps (NETs) bind to bacteria, prevent them from spreading, and kill them with antimicrobial proteins such as histones and elastins. One hypothesis, shown in this animation, proposes that the NETs are produced by living neutrophils in response to bacteria. Alternately, NETs may be released as a result of necrotic cell death of neutrophils.
Some microorganisms are more resistant to phagocytic attachment.
a. Capsules can resist unenhanced attachment by preventing the endocytic pattern recognition receptors on phagocytes from recognizing the bacterial cell wall components and mannose-containing carbohydrates (see Figure \(\PageIndex{14}\)). Streptococcus. pneumonia activates the classical complement pathway, but resists C3b opsonization, and complement causes further inflammation in the lungs.
Movie of an encapsulated bacterium resisting engulfment by a neutrophil. Phagocytosis. © James Sullivan, author. Licensed for use, ASM MicrobeLibrary.
b. Some capsules prevent the formation of C3 convertase, an early enzyme in the complement pathways. Without this enzyme, the opsonins C3b and C4b, as well as the other beneficial proteins are not produced.
c. Other capsules, rich in sialic acid, a common component of host cell glycoprotein, have an affinity for serum protein H, a complement regulatory protein that leads to the degradation of the opsonin C3b by factor I and the formation of C3 convertase. (Serum protein H is what normally leads to the degradation of any C3b that binds to host glycoproteins so that we don't stick our own phagocytes to our own cells with C3b.)
d. Some capsules simply cover the C3b that does bind to the bacterial surface and prevent the C3b receptor on phagocytes from making contact with the C3b (see Figure \(\PageIndex{3}\)). This is seen with the capsule of Streptococcus pneumoniae.
e. Neisseria meningitidis has a capsule composed of sialic acid while Streptococcus pyogenes (group A beta streptococci) has a capsule made of hyaluronic acid. Both of these polysaccharides closely resemble carbohydrates found in human tissue polysaccharides and because they are not recognized as foreign by the lymphocytes that carry out the immune responses, antibodies are not made against these capsules. Likewise, some bacteria are able to coat themselves with host proteins such as fibronectin, lactoferrin, or transferrin and in this way avoid antibodies.
f. An outer membrane molecule of Neisseria gonorrhoeae called Protein II and the M-protein of Streptococcus pyogenes allow these bacteria to be more resistant to phagocytic engulfment. The M-protein of S. pyogenes, for example, binds factor H of the complement pathway and this results in the degradation of the opsonin C3b by factor I and the formation of C3 convertase. S. pyogenes also produces a protease that cleaves the complement protein C5a.
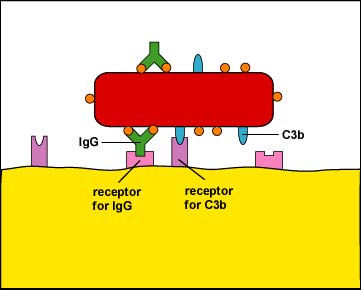
g. Staphylococcus aureus produces protein A while Streptococcus pyogenes produces protein G. Both of these proteins bind to the Fc portion of antibodies (see Figure \(\PageIndex{4}\)) and in this way the bacteria become coated with antibodies in a way that does not result in opsonization (see Figure \(\PageIndex{5}\)).
Step 4: Ingestion of the Microbe or Cell by the Phagocyte
Following attachment, polymerization and then depolymerization of actin filaments send pseudopods out to engulf the microbe (see Figure \(\PageIndex{6}\)) and place it in an endocytic vesicle called a phagosome (see Figure \(\PageIndex{7}\)).
During this process, an electron pump brings protons (H+) into the phagosome. This lowers the pH within the phagosome to 3.5 - 4.0 so that when a lysosome fuses with the phagosome, the pH is correct for the acid hydrolases to effectively break down cellular proteins. The acidification also releases defensins, cathelicidin, and bacterial permeability inducing protein (BPI), peptides and enzymes that can kill microbes, from a matrix and enabling their activation.
Scanning electron micrographs of a macrophage with pseudopods and a macrophage phagocytozing E. coli on a blood vessel; courtesy of Dennis Kunkel's Microscopy.
Intracellular microbes, such as viruses and bacteria that invade host cells, can also be engulfed once they enter the cytosol of the cell by a process called autophagy. A membrane-bound compartment called an autophagosome grows around the microbe and the surrounding cytosol and subsequently delivers it to lysosomes for destruction (see Figure \(\PageIndex{17}\)). (This process is also used by eukaryotic cells to engulf and degrade unnecessary or dysfunctional cellular components such as damaged organelles.)
Some microorganisms are more resistant to phagocytic ingestion
a. Pathogenic Yersinia, such as the one that causes plague, contact phagocytes and, by means of a type III secretion system, deliver proteins which depolymerize the actin microfilaments needed for phagocytic engulfment into the phagocytes (see Figure \(\PageIndex{8}\)). Another Yersinia protein degrades C3b and C5a.
b. Some bacteria, like Mycobacterium tuberculosis, Salmonella, and Listeria monocytogenes can block autophagy.
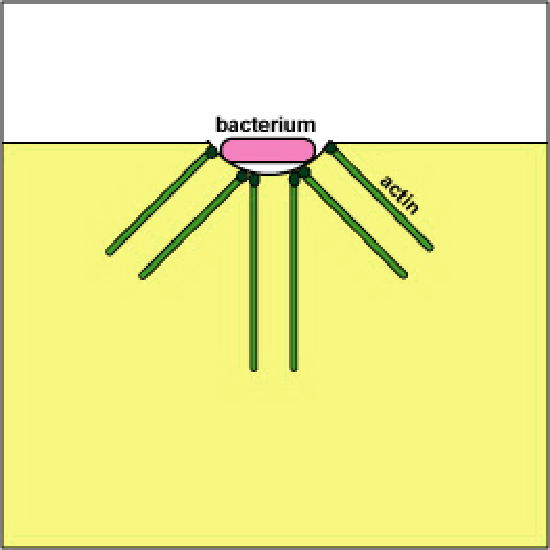
Blocking Phagosome Formation by Depolymerizing Actin. Molecules of some bacteria, through a type III secretion system, deliver proteins which depolymerize the phagocyte's actin microfilaments used for phagocytic engulfment.
Step 5: Destruction of the Microbe or CellPhagocytes contain membranous sacs called lysosomes produced by the Golgi apparatus that contain various digestive enzymes, microbicidal chemicals, and toxic oxygen radicals. The lysosomes travel along microtubules within the phagocyte and fuse with the phagosomes containing the ingested microbes and the microbes are destroyed (see Figure \(\PageIndex{9}\)).
To view an electron micrograph of a phagolysosome, see the Web page for the University of Illinois College of Medicine.
| You Tube animation summarizing phagocytosis by a macrophage. |
Some bacteria are more resistant to phagocytic destruction once engulfed.
a. Some bacteria, such as Legionella pneumophilia and Mycobacterium species, cause the phagocytic cell to place them into an endocytic vacuole via a pathway that decreases their exposure to toxic oxygen compounds.
b. Some bacteria, such as Salmonella, are more resistant to toxic forms of oxygen and to defensins (toxic peptides that kill bacteria).
c. Some bacteria, such as Shigella flexneri and the spotted fever Rickettsia, escape from the phagosome into the cytoplasm prior to the phagosome fusing with a lysosome (see Figure \(\PageIndex{10}\)).
d. Neisseria gonorrhoeae produces Por protein (protein I) that prevents phagosomes from fusing with lysosomes enabling the bacteria to survive inside phagocytes.
e. Some bacteria, such as species of Salmonella, Mycobacterium, Legionella, and Chlamydia, block the vesicular transport machinery that enables the phagosome to fuse with the lysosome.
f. Some bacteria, such as pathogenic Mycobacterium and Legionella pneumophilia, prevent the acidification of the phagosome which is needed for effective killing of microbes by lysosomal enzymes. (Normally after the phagosome forms, the contents become acidified because the lysosomal enzymes used for killing function much more effectively at an acidic pH.)
g. The carotenoid pigments that give Staphylococcus aureus its golden color and group B streptococci (GBS) its orange tint shield the bacteria from the toxic oxidants that neutrophils use to kill bacteria.
h. Cell wall lipids of Mycobacterium tuberculosis, such as lipoarabinomannan, arrest the maturation of phagosomes preventing delivery of the bacteria to lysosomes.
i. Some bacteria are able to kill phagocytes. Bacteria such as Staphylococcus aureus and Streptococcus pyogenes produce the exotoxin leukocidin which damages the cytoplasmic membrane of the phagocyte. On the other hand, bacteria, such as Shigella and Salmonella, induce macrophage apoptosis, a programmed cell death.
If the the infection site contains very large numbers of microorganisms and high levels of inflammatory cytokines and chemokines are being produced in response to PAMPs, the phagocyte will empty the contents of its lysosomes by a process called degranulation in order to kill the microorganisms or cell extracellularly.
These released lysosomal contents, however, also kill surrounding host cells and tissue. Most tissue destruction associated with infections is a result of this process (see Figure \(\PageIndex{11}\)).
The phagocyte will also empty the contents of its lysosomes for extracellular killing if the cell to which the phagocyte adheres is too large to be engulfed (see Figure \(\PageIndex{12}\) and Figure \(\PageIndex{13}\)).
There are 2 killing systems in neutrophils and macrophages: the oxygen-dependent system and the oxygen-independent system.
1. The oxygen-dependent system: production of reactive oxygen species (ROS)
The cytoplasmic membrane of phagocytes contains the enzyme oxidase which converts oxygen into superoxide anion (O2-). This can combine with water by way of the enzyme dismutase to form hydrogen peroxide (H2O2) and hydroxyl (OH) radicals.
In the case of neutrophils, but not macrophages, the hydrogen peroxide can then combine with chloride (Cl2-) ions by the action of the enzyme myeloperoxidase (MPO) to form hypochlorous acid (HOCL), and singlet oxygen.
In macrophages, nitric oxide (NO) can combine with hydrogen peroxide to form peroxynitrite radicals. (In addition to ROS and NO, macrophages secrete inflammatory cytokines such as TNF-alpha, IL-1, IL-8, and IL-12 to promote an inflammatory response.)
These compounds are very microbicidal because they are powerful oxidizing agents which oxidize most of the chemical groups found in proteins, enzymes, carbohydrates, DNA, and lipids. Lipid oxidation can break down cytoplasmic membranes. Collectively, these oxidizing free radicals are called reactive oxygen species (ROS).
Oxidase also acts as an electron pump that brings protons (H+) into the phagosome. This lowers the pH within the phagosome so that when lysosomes fuse with the phagosome, the pH is correct for the acid hydrolases, like elastase, to effectively break down cellular proteins.
In addition to phagocytes using this oxygen-dependant system to kill microbes intracellularly, neutrophils also routinely release these oxidizing agents, as well as acid hydrolases, for the purpose of killing microbes extracellularly. These agents, however, also wind up killing the neutrophils themselves as well as some surrounding body cells and tissues as mentioned above.
2. The oxygen-independent system
Some lysosomes contain defensins ), cationic peptides that alter cytoplasmic membranes; lysozyme, an enzyme that breaks down peptidoglycan, lactoferrin, a protein that deprives bacteria of needed iron; cathepsin G, a protease that causes damage to microbial membranes; elastase, a protease that kills many types of bacteria; cathelicidins, proteins that upon cleavage are directly toxic to a variety of microorganisms; bactericidal permeability inducing protein (BPI ), proteins used by neutrophils to kill certain bacteria by damaging their membranes; collagenase ; and various other digestive enzymes that exhibit antimicrobial activity by breaking down proteins, RNA, phosphate compounds, lipids, and carbohydrates.
Streptococcus pyogenes has a capsule made of hyaluronic acid, a polysaccharide also found on human cells.
Describe what significance this has in the bacterium resisting phagocytosis and why.
- Streptococcus pyogenes produces a protein called protein G that binds to the Fc portion of human IgG.
Describe what significance this has in the bacterium resisting phagocytosis and why.
- Many bacteria produces capsules that cover their cell wall.
Describe what significance this has in the bacterium resisting phagocytosis and why.
- People born with chronic granulomatious disease have neutrophils that lack the enzyme oxidase in their cytoplasmic membrane.
Describe what significance this has in the bacterium resisting phagocytosis and why.
Summary
Phagocytosis is the primary method used by the body to remove free microorganisms in the blood and tissue fluids. An inflammatory response to injury and/or infection allows phagocytes to leave the bloodstream, enter the tissue, and go to the site of infection or injury. Microorganisms entering lymph nodules found in the respiratory, gastrointestinal, and genitourinary tract can be phagocytosed by fixed macrophages and dendritic cells and presented to B-lymphocytes and T-lymphocytes to initiate adaptive immune responses.Tissue fluid picks up microbes in the tissue, enters the lymph vessels as lymph, and carries the microbes to regional lymph nodes where they are filtered out and phagocytosed by fixed macrophages and dendritic cells and presented to the circulating B-lymphocytes and T-lymphocytes to initiate adaptive immune responses.
Dendritic cells located throughout the epithelium of the skin, the respiratory tract, and the gastrointestinal tract phagocytize microbes, enter lymph vessels, and carry the microbes to regional lymph nodes where the dendritic cells present antigens associated with the microbes to the ever changing populations of naive T-lymphocytes.Blood carries microorganisms to the spleen where they are filtered out and phagocytosed by fixed macrophages and dendritic cells and presented to the circulating B-lymphocytes and T-lymphocytes to initiate adaptive immune responses. There are also specialized macrophages and dendritic cells located in the brain (microglia), lungs (alveolar macrophages), liver (Kupffer cells), kidneys (mesangial cells), bones (osteoclasts), and the gastrointestinal tract (peritoneal macrophages.
- Resting phagocytes are activated by inflammatory mediators and produce surface receptors that increase their ability to adhere to the inner surface of capillary walls enabling them to squeeze out of the capillary and enter the tissue, a process called diapedesis.
- Activation also enables phagocytes to produce endocytic pattern-recognition receptors that recognize and bind to microbial PAMPs in order to attach the microbe to the phagocyte, as well as to exhibit increased metabolic and microbicidal activity.
- Phagocytes then use chemotaxis to move towards an increasing concentration of some attractant such as bacterial factors or defense molecules.
- Attachment of phagocytes to the microbes or cells can be through unenhanced attachment or enhanced attachment.
- Unenhanced attachment is the recognition of pathogen-associated molecular patterns or PAMPs by endocytic pattern-recognition receptors on the surface of the phagocytes.
- Enhanced attachment, or opsonization, is the attachment of microbes to phagocytes by way of an antibody molecule called IgG, the complement proteins C3b and C4b, and acute phase proteins such as mannose-binding lectin (MBL) and C-reactive protein (CRP).
- Following attachment, polymerization and then depolymerization of actin filaments send pseudopods out to engulf the microbe and place it in an endocytic vesicle called a phagosome.
- During this process, an electron pump brings protons (H+) into the phagosome to lowers the pH within the phagosome to a pH that is correct for the acid hydrolases to effectively break down cellular proteins.
- Phagocytes contain membranous sacs called lysosomes that contain various digestive enzymes, microbicidal chemicals, and toxic oxygen radicals. The lysosomes fuse with the phagosomes containing the ingested microbes and the microbes are destroyed.
- If the infection site contains very large numbers of microorganisms and high levels of inflammatory cytokines and chemokines are being produced in response to PAMPs, the phagocyte will empty the contents of its lysosomes in order to kill the microorganisms or cell extracellularly.
- Lysosomal contents released during extracellular killing also kill surrounding host cells and tissue. Most tissue destruction associated with infections is a result of extracellular killing by phagocytes.