
11.3B: The Complement System
- Page ID
- 3271

Briefly describe how the classical complement pathway is activated. Briefly describe the beneficial effects of the following complement pathway products: - C5a
- C3a
- C3b
- C4b
- C3d
- C5b6789n (MAC)
Briefly describe how the lectin pathway is activated. Briefly describe how the alternative complement pathway is activated.
- C5a
- C3a
- C3b
- C4b
- C3d
- C5b6789n (MAC)
In this section we will look at how the body's complement system functions to remove infectious agents. The complement system refers to a series of more than 30 soluble, preformed proteins circulating in the blood and bathing the fluids surrounding tissues. The proteins circulate in an inactive form, but in response to the recognition of molecular components of microorganism, they become sequentially activated, working in a cascade where in the binding of one protein promotes the binding of the next protein in the cascade. There are 3 complement pathways that make up the complement system: the classical complement pathway, the lectin pathway, and the alternative complement pathway. The pathways differ in the manner in which they are initiated and ultimately produce a key enzyme called C3 convertase:
- The classical complement pathway is initiated by activation of C1. C1 is primarily activated by interacting with the Fc portion of the antibody molecules IgG or IgM after they have bound to their specific antigen. C1 is also able to directly bind to the surfaces of some pathogens as well as with the C-reactive protein (CRP) that is produced during the acute phase response of innate immunity.
- The lectin pathway is activated by the interaction of microbial carbohydrates (lectins) with mannose-binding lectin (MBL) or ficolins found in the plasma and tissue fluids.
- The alternative complement pathway is activated by C3b binding to microbial surfaces and to antibody molecules.
The end results and defense benefits of each pathway, however, are the same. All complement pathways carry out 6 beneficial innate defense functions. Proteins produced by the complement pathways:
- Trigger inflammation,
- Chemotactically attract phagocytes to the infection site,
- Promote the attachment of antigens to phagocytes (enhanced attachment or opsonization),
- Cause lysis of Gram-negative bacteria, human cells displaying foreign epitopes,and viral envelopes,
- Play a role in the activation of naive B-lymphocytes during adaptive immunity, and
- Remove harmful immune complexes from the body.
We will now look at each of these complement pathways and see how they function to protect the body.
The Classical Complement Pathway
The classical complement pathway is primarily activated when a complement protein complex called C1 interacts with the Fc portion of the antibody molecules IgG or IgM after they have bound to their specific antigen via their Fab portion. C1 is also able to directly bind to the surfaces of some pathogens as well as with the C-reactive protein (CRP) that is produced during the acute phase response of innate immunity. The C1 complex is composed of three complement proteins called C1q, C1r, and C1s.
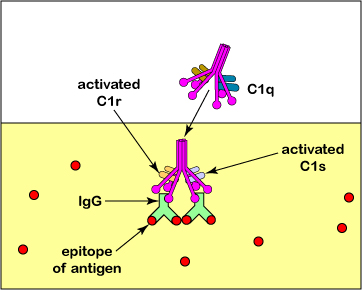
C1 is also able to directly bind to the surfaces of some pathogens as well as with the C-reactive protein (CRP) that is produced during the acute phase response of innate immunity.
2. The binding of C1q activates the C1r portion of C1 which, in turn, activates C1s. This activation gives C1s enzymatic activity to cleave complement protein C4 into C4a and C4b (see Figure \(\PageIndex{2}\)A and Figure \(\PageIndex{2}\)B).
3. C2 then binds to C4b and is cleaved by C1 into C2a and C2b (see Figure \(\PageIndex{3}\)A and Figure \(\PageIndex{3}\)B).
4. C4b and C2a combine to form C4b2a, the C3 convertase. C3 convertase can now cleave hundreds of molecules of C3 into C3a and C3b (see Figure \(\PageIndex{4}\)).
5. Some molecules of C3b bind to C4b2a, the C3 convertase, to form C4b2a3b, a C5 convertase that cleaves C5 into C5a and C5b (see Figure \(\PageIndex{5}\)).
6. C5b binds to the surface of the target cell and subsequently binds C6, C7, C8, and a number of monomers of C9 to form C5b6789n, the Membrane Attack Complex (MAC) (see Figure \(\PageIndex{6}\) and Figure \(\PageIndex{7}\)).
As mentioned above, proteins of the complement pathways carry out 6 beneficial innate defense functions. These include:
1. Triggering inflammation: C5a is the most potent complement protein triggering inflammation. It reacts with blood vessels causing vasodilation. It also causes mast cells to release vasodilators such as histamine,increasing blood vessel permeability as well as increasing the expression of adhesion molecules on leukocytes and the vascular endothelium so that leukocytes can squeeze out of the blood vessels and enter the tissue (diapedesis). C5a also causes neutrophils to release toxic oxygen radicals for extracellular killing and induces fever. To a lesser extent C3a and C4a also promote inflammation. As we will see later in this unit, inflammation is a process in which blood vessels dilate and become more permeable, thus enabling body defense cells and defense chemicals to leave the blood and enter the tissues.
2. Chemotactically attracting phagocytes to the infection site: C5a also functions as a chemoattractant for phagocytes. Phagocytes will move towards increasing concentrations of C5a and subsequently attach, via their CR1 receptors to the C3b molecules attached to the antigen. This will be discussed in greater detail later in this unit under phagocytosis.
3. Promoting the attachment of antigens to phagocytes (enhanced attachment or opsonization): C3b and to a lesser extent, C4b can function as opsonins, that is, they can attach antigens to phagocytes. One portion of the C3b binds to proteins and polysaccharides on microbial surfaces; another portion attaches to CR1 receptors on phagocytes, B-lymphocytes, and dendritic cells for enhanced phagocytosis. (see Figure \(\PageIndex{8}\)). In actuality, C3b molecule can bind to pretty much any protein or polysaccharide. Human cells, however, produce Factor H that binds to C3b and allows Factor I to inactivate the C3b. On the other hand, substances such as LPS on bacterial cells facilitate the binding of Factor B to C3b and this protects the C3b from inactivation by Factor I. In this way, C3b does not interact with our own cells but is able to interact with microbial cells. C3a and C5a increase the expression of C3b receptors on phagocytes and increase their metabolic activity.
4. Causing lysis of Gram-negative bacteria, human cells displaying foreign epitopes,and viral envelopes: C5b6789n, functions as a Membrane Attack Complex (MAC). This helps to destroy gram-negative bacteria as well as human cells displaying foreign antigens (virus-infected cells, tumor cells, etc.) by causing their lysis; see Figure \(\PageIndex{6}\) and Figure \(\PageIndex{7}\). It can also damage the envelope of enveloped viruses.
5. Serving as a second signal for activating naive B-lymphocytes during adaptive immunity: Some C3b is converted to C3d. C3d binds to CR2 receptors on B-lymphocytes. This serves as a second signal for the activation of B-lymphocytes whose B-cell receptors have just interacted with their corresponding antigen.
6. Removing harmful immune complexes from the body: C3b and to a lesser extent, C4b help to remove harmful immune complexes from the body. The C3b and C4b attach the immune complexes to CR1 receptors on erythrocytes. The erythrocytes then deliver the complexes to fixed macrophages within the spleen and liver for destruction. Immune complexes can lead to a harmful Type III hypersensitivity, as will be discussed later in Unit 5 under Hypersensitivities.
Some bacterial capsules are rich in sialic acid, a common component of host cell glycoprotein, that has an affinity for serum protein H, a complement regulatory protein that leads to the degradation of C3b.
Describe what significance this has in the bacterium resisting phagocytosis and why.
- S. pyogenes produces a protease that cleaves the complement protein C5a.
Describe what significance this has in the bacterium resisting phagocytosis and why.
The Lectin Pathway
The lectin pathway is activated by the interaction of microbial carbohydrates with mannose-binding lectin (MBL) or ficolins found in the plasma and tissue fluids. (Lectins are carbohydrate-binding proteins.) The lectin pathway is mediated by two groups of proteins found in the plasma of the blood and in tissue fluids:
1. Mannose-binding lectin (MBL) - also known as mannose-binding protein or MBP. MBL is a soluble pattern-recognition receptor that binds to various microbial carbohydrates such as those rich in mannose or fucose, and to N-acetylglucosamine (NAG). These glycans are common in microbial glycoproteins and glycolipids but rare in those of humans. MBL is synthesized by the liver and released into the bloodstream as part of the acute phase response that will be discussed later in this unit. The MBL is equivalent to C1q in the classical complement pathway.
Ficolins are similar in their structure to MBL and bind to microbial carbohydrates such as N-acetylglucosamine (NAG), lipoteichoic acids, and lipopolysaccharide (LPS). Ficolin is also equivalent to C1q in the classical complement pathway.
2. Both mannose-binding lectin (MBL) and ficolin form complexes with MBL-associated serine proteases called MASP1 and MASP2, which are equivalent to C1r and C1s of the classical pathway.
a. The binding of the MBL (or the ficolin) to the microbial carbohydrate activates the associated MASP2 giving it the enzymatic activity to split C4 into C4a and C4b (see Figure \(\PageIndex{9}\)A and Figure \(\PageIndex{9}\)B).
b. C2 then binds to C4b and is cleaved by MASP2 into C2a and C2b (see Figure \(\PageIndex{10}\)A and Figure \(\PageIndex{10}\)B).
c. C4b and C2a combine to form C4b2a, the C3 convertase. C3 convertase can now cleave hundreds of molecules of C3 into C3a and C3b (see Figure \(\PageIndex{11}\)).
d. Some molecules of C3b bind to C4b2a, the C3 convertase, to form C4b2a3b, a C5 convertase that cleaves C5 into C5a and C5b (see Figure \(\PageIndex{12}\)).
e. C5b binds to the surface of the target cell and subsequently binds C6, C7, C8, and a number of monomers of C9 to form C5b6789n, the Membrane Attack Complex (MAC) (see Figure \(\PageIndex{6}\) and Figure \(\PageIndex{7}\)).
The beneficial results of the activated complement proteins are the same as in the classical complement pathway above. The complement proteins:
1. Trigger inflammation : C5a>C3a>c4a;
2. Chemotactically attract phagocytes to the infection site: C5a;
3. Promote the attachment of antigens to phagocytes via enhanced attachment or opsonization : C3b>C4b;
4. Cause lysis of Gram-negative bacteria and human cells displaying foreign epitopes : MAC;
5. Serve as a second signal for the activation of naive B-lymphocytes ): C3d; and
6 Remove harmful immune complexes from the body: C3b>C4b.
The Alternative Complement Pathway
The alternative complement pathway is mediated by C3b, produced either by the classical or lectin pathways or from C3 hydrolysis by water. (Water can hydrolyze C3 and form C3i, a molecule that functions in a manner similar to C3b.)
Activation of the alternative complement pathway begins when C3b (or C3i) binds to the cell wall and other surface components of microbes. C3b can also bind to IgG antibodies. Alternative pathway protein Factor B then combines with the cell-bound C3b to form C3bB. Factor D then splits the bound Factor B into Bb and Ba, forming C3bBb. A serum protein called properdin then binds to the Bb to form C3bBbP that functions as a C3 convertase (see Figure \(\PageIndex{13}\)) capable of enzymatically splitting hundreds of molecules of C3 into C3a and C3b. The alternative complement pathway is now activated.
Some of the C3b subsequently binds to some of the C3bBb to form C3bBb3b, a C5 convertase capable of splitting molecules of C5 into C5a and C5b (see Figure \(\PageIndex{14}\)). From here, the alternative complement pathway is identical to the other complement pathways.
The beneficial results are the same as in the classical complement pathway above. The complement proteins:
- Trigger inflammation : C5a>C3a>c4a;
- Chemotactically attract phagocytes to the infection site: C5a;
- Promote the attachment of antigens to phagocytes via enhanced attachment or opsonization : C3b>C4b;
- Cause lysis of Gram-negative bacteria, human cells displaying foreign epitopes,and viral envelopes: MAC; and
- Serve as a second signal for the activation of naive B-lymphocytes ): C3d;
- Remove harmful immune complexes from the body: C3b>C4b.
Keep in mind that in Unit 3, we learned several mechanisms that various bacteria use to resist the body's complement pathways. By resisting these immediate innate immune defenses, some bacteria have a better chance of colonizing their host.
Summary
- The proteins of the complement system circulate in an inactive form, but in response to the recognition of molecular components of microorganism, they become sequentially activated, working in a cascade where in the binding of one protein promotes the binding of the next protein in the cascade.
- There are 3 complement pathways that make up the complement system: the classical complement pathway, the lectin pathway, and the alternative complement pathway.
- The classical complement pathway is initiated by activation of C1. C1 is primarily activated by interacting with the Fc portion of the antibody molecules IgG or IgM after they have bound to their specific antigen. C1 is also able to directly bind to the surfaces of some pathogens as well as with the C-reactive protein (CRP) that is produced during the acute phase response of innate immunity.
- The lectin pathway is activated by the interaction of microbial carbohydrates (lectins) with mannose-binding lectin (MBL) or ficolins found in the plasma and tissue fluids.
- The alternative complement pathway is activated by C3b binding to microbial surfaces and to antibody molecules.
- All complement pathways carry out the same 6 beneficial innate defense functions.
- The complement proteins C5a and, to a lesser extent, C3a, and C4a trigger vasodilation and inflammation in order to deliver defense cells and defense chemicals to the infection site.
- The complement protein C5a also functions as a chemoattractant for phagocytes.
- The complement proteins C3b and to a lesser extent, C4b can function as opsonins, that is, they can attach antigens to phagocytes.
- The complement proteins C5b6789n, functions as a Membrane Attack Complex (MAC) causing lysis of Gram-negative bacteria, human cells displaying foreign epitopes, and viral envelopes.
- The complement protein C3d serves as a second signal for activating naive B-lymphocytes during adaptive immunity.
- The complement proteins C3b and to a lesser extent, C4b help to remove harmful immune complexes from the body.