
16.3E: Self-incompatibility - How Plants Avoid Inbreeding
- Page ID
- 5793

Evolution seems to favor (and be favored by) genetic variability. Genetic variability is promoted by outbreeding - sexual reproduction between genetically dissimilar parents. Just why sexual reproduction is so popular throughout the world of living things is still a hotly-debated question, but the fact remains. Plants, being anchored in position, have a special problem in this regard. Many employ the services of animals (e.g., insects, birds, bats) to transfer pollen from plant to plant. But if the flowers have both sex organs:what is to prevent the pollen from fertilizing its own eggs?
A variety of solutions have been tried in the plant kingdom. These include:
- Having imperfect flowers; that is, flowers that are either male or female.
- Dioecy. The imperfect flowers are present on separate plants. Dioecy is the equivalent of the separate sexes of most animals. But it is rather rare. Some examples include poplars and hollies.
- Monoecy. The imperfect flowers are present on the same plant. But if they mature at different times, self-fertilization is avoided. Corn (maize) is a common example.
But the vast majority of angiosperms have perfect flowers; that is containing both male and female sex organs. So how do they avoid self-fertilization?
- Heteromorphic flowers.
The flowers are perfect but come in two structural types; for example
- long stamens with a short style
- short stamens with a long style
- Homomorphic flowers. All flowers have exactly the same structure. Avoidance of self-fertilization depends on genetic/biochemical mechanisms. There are two quite different types of self-incompatibility.
- Sporophytic self-incompatibility (SSI)
- Gametophytic self-incompatibility (GSI)
Sporophytic Self-Incompatibility (SSI)
This form of self-incompatibility has been studied intensively in members of the mustard family (Brassica), including turnips, rape, cabbage, broccoli, and cauliflower.
In this system,
- Rejection of self pollen is controlled by the diploid genotype of the sporophyte generation.
- The control lies in the "S-locus", which is actually a cluster of three tightly-linked loci:
- SLG (S-Locus Glycoprotein) which encodes part of a receptor present in the cell wall of the stigma;
- SRK (S-Receptor Kinase), which encodes the other part of the receptor. Kinases attach phosphate groups to other proteins. SRK is transmembrane protein embedded in the plasma membrane of the stigma cell.
- SCR (S-locus Cysteine-Rich protein), which encodes a soluble ligand for the same receptor which is secreted by the pollen.
- Because the plants cannot fertilize themselves, they tend to be heterozygous; that is, carry a pair of different S loci (here designated S1 and S2).
- However, dozens of different S alleles may be present in the population of the species; that is; the S-locus in the species is extremely polymorphic (analogous to the major histocompatibility locus of vertebrates).
- The difference between the alleles is concentrated in certain "hypervariable regions" of the receptor (analogous to the hypervariable regions that provide the great binding diversity of antibodies

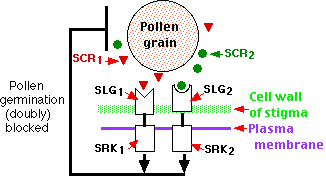
The rules:
- Pollen will not germinate on the stigma (diploid) of a flower that contains either of the two alleles in the sporophyte parent that produced the pollen.
- This holds true even though each pollen grain being haploid contains only one of the alleles.
- In the example shown here, the S2 pollen, which was produced by a S1S2 parent, cannot germinate on an S1S3 stigma.
The explanation:
- The S1S2 pollen-producing sporophyte synthesizes both SCR1 and SCR2 for incorporation in (and later release from) both S1 and S2 pollen grains.
- If either SCR molecule can bind to either receptor on the pistil, the kinase triggers a series of events that lead to failure of the stigma to support germination of the pollen grain. Among these events is the ubiquination of proteins targeting them for destruction in proteasomes.
- If this path is not triggered (e.g., pollen from an S1S2 parent on an S3S4 stigma, the pollen germinates successfully.
Gametophytic Self-Incompatibility (GSI)
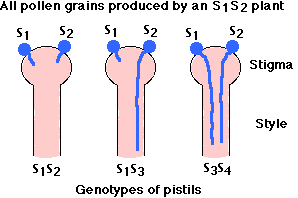
This form of self-incompatibility is more common than SSI but not so well understood. It occurs in nearly one-half of all the families of angiosperms, including
- the Solanaceae (potatoes, tomatoes [wild, not cultivated], and tobacco)
- petunias
- beets (Beta vulgaris)
- buttercups (Ranunculus)
- lilies
- roses
- many grasses
The rules:
- The S loci are (as in SSI plants) extremely polymorphic; that is, there is an abundance of multiple alleles in the population.
- Incompatibility is controlled by the single S allele in the haploid pollen grain.
- Thus a pollen grain will grow in any pistil that does not contain the same allele (so, as shown here and in contrast to what happens in SSI, S2 pollen from an S1S2 parent will grow down an S1S3 style.
This appears to be the mechanism in the petunia:
- All pollen grains - incompatible as well as compatible - germinate forming pollen tubes that begin to grow down the style.
- However, growth of incompatible pollen tubes stops in the style while compatible tubes go on to fertilize the egg in the ovary.
- The block within incompatible pollen tubes is created by an S-locus-encoded ribonuclease (S-RNase), which is synthesized within the style, enters the pollen tube and destroys its RNA molecules halting pollen tube growth.
- The RNase molecules contain a hypervariable region, each encoded by a different allele, which establishes each S specificity (S1, S2, S3, etc.).
- The pollen tube expresses a protein designated SLF that binds S-RNase. SLF also exists in different S specificities (S1, S2, S3, etc.).
- In compatible ("nonself") tubes, the SCF triggers the degradation (in proteasomes) of the S-RNase thus permitting RNAs in the pollen tube to survive and growth to continue.
- In incompatible ("self") tubes the interaction of, for example, the S1 SCF with the S1 S-RNase blocks its degradation so the RNAs of the pollen tube are destroyed and growth is halted.
An entirely-different mechanism of gametophytic self-incompatibility is found in poppies (Papaver rhoeas).
Switching from Cross-Pollination to Self-Pollination
A substantial minority of angiosperms have abandoned cross-pollination for self-pollination. For example, while its wild relatives continue to be cross-pollinated, the domestic tomato is not. Two steps are needed for this change:
- abandoning its mechanism of self-incompatibility
- changes in flower structure to reduce the chance that pollinators will transfer pollen from another plant to its stigma.
Unlike its wild relatives, the stigma of the domestic tomato does not protrude beyond the anthers. Of the several genes involved in this change, the most important one is Style2.1. The mutation in Style2.1 responsible for the change in phenotype in our cultivated tomatoes is found in the promoter region - the protein-encoding portion of the gene is exactly the same as in wild tomatoes.
Here, again, is evidence that much of the diversity of life arises not from mutations in the protein-coding portion of the genes that we share but mutations in their regulatory regions (promoters and enhancers).
Contributors and Attributions
John W. Kimball. This content is distributed under a Creative Commons Attribution 3.0 Unported (CC BY 3.0) license and made possible by funding from The Saylor Foundation.