
3.7: Centrosomes and Centrioles
- Page ID
- 3971

Centrioles
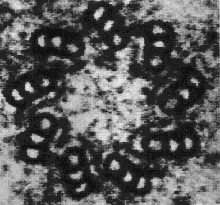
Centrioles are built from a cylindrical array of 9 microtubules, each of which has attached to it 2 partial microtubules. Figure \(\PageIndex{1}\) is an electron micrograph showing a cross section of a centriole with its array of nine triplets of microtubules.
When a cell enters the cell cycle and passes through S phase, each centriole is duplicated. A "daughter" centriole grows out of the side of each parent ("mother") centriole. Thus centriole replication — like DNA replication (which is occurring at the same time) — is semiconservative.
- Functional microtubules grow out only from the "mother".
- When stem cells divide, one daughter cell remains a stem cell; the other goes on to differentiate. In two animal systems that have been examined (mouse glial cells and Drosophila male germline cells), the cell that receives the old ("mother") centriole remains a stem cell while the one that receives what had been the original "daughter" centriole goes on to differentiate. (You can read about these findings in Wang, X., et. al., Nature, 15 October 2009.)
Centrioles are a key feature of eukaryotic cells and presumably arose with the first eukaryotes. A few groups have since lost their centrioles including most fungi (but not the primitive chytrids), "higher" plants (but not the more primitive mosses, ferns, and cycads with their motile sperm) and animal eggs lose their centriole during meiosis and must have it restored by the sperm that fertilizes it
In nondividing cells, the mother centriole can attach to the inner side of the plasma membrane forming a basal body. In almost all types of cell, the basal body forms a nonmotile primary cilium. In cells with a flagellum, e.g. sperm, the flagellum develops from a single basal body. (While sperm cells have a basal body, eggs have none. So the sperm's basal body is absolutely essential for forming a centrosome which will form a spindle enabling the first division of the zygote to take place.)
In ciliated cells such as the columnar epithelial cells of the lungs and ciliated protozoans like the paramecium, many basal bodies form, each producing a beating cilium. Most of their centrioles are produced by repeated duplication of the daughter centriole of centrosome and are temporarily assembled in a special organelle called the deuterosome (not to be confused with deuterostome). Centrioles organize the centrosome in which they are embedded.
Centrosomes
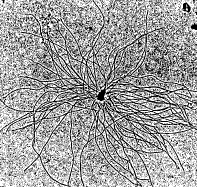
The centrosome is located in the cytoplasm usually close to the nucleus. It consists of two centrioles — oriented at right angles to each other — embedded in a mass of amorphous material containing more than 100 different proteins.It is duplicated during S phase of the cell cycle. Just before mitosis, the two centrosomes move apart until they are on opposite sides of the nucleus. As mitosis proceeds, microtubules grow out from each centrosome with their plus ends growing toward the metaphase plate. These clusters of microtubules are called spindle fibers. Figure \(\PageIndex{2}\) shows microtubules growing in-vitro from an isolated centrosome. The centrosome was supplied with a mixture of alpha and beta tubulin monomers spontaneously assembled into microtubules only in the presence of centrosomes.
Spindle fibers have three destinations:
- Some attach to one kinetochore of a dyad with those growing from the opposite centrosome binding to the other kinetochore of that dyad.
- Some bind to the arms of the chromosomes.
- Still others continue growing from the two centrosomes until they extend between each other in a region of overlap.
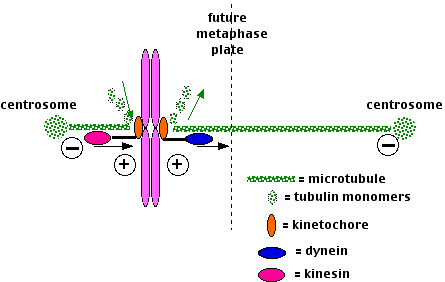
All three groups of spindle fibers participate in
- the assembly of the chromosomes at the metaphase plate at metaphase.
- Microtubules attached to opposite sides of the dyad shrink or grow until they are of equal length.
- Microtubules motors attached to the kinetochores move them
- toward the minus end of shrinking microtubules (a dynein);
- toward the plus end of lengthening microtubules (a kinesin).
- The chromosome arms use a different kinesin to move to the metaphase plate.
- the separation of the chromosomes at anaphase.
- The sister kinetochores separate and, carrying their attached chromatid,
- move along the microtubules powered by minus-end motors, dyneins, while the microtubules themselves shorten (probably at both ends).
- The overlapping spindle fibers move past each other (pushing the poles farther apart) powered by plus-end motors, the "bipolar" kinesins.
- In this way the sister chromatids end up at opposite poles.
Functions of Centrosomes
In addition to their role in spindle formation, centrosomes play other important roles in animal cells:
- Formation of the network of microtubules that participate in making the cytoskeleton.
- Signaling that it is o.k. to proceed to cytokinesis. Destruction of both centrosomes with a laser beam prevents cytokinesis even if mitosis has been completed normally.
- Signaling that it is o.k. for the daughter cells to begin another round of the cell cycle; specifically to duplicate their chromosomes in the next S phase. Destruction of one centrosome with a laser beam still permits cytokinesis but the daughter cells fail to enter a new S phase.
- Segregating signaling molecules (e.g., mRNAs) so that they pass into only one of the two daughter cells produced by mitosis. In this way, the two daughter cells can enter different pathways of differentiation even though they contain identical genomes.
- In at least some developing neurons, the position of the centrosome establishes the point at which the axon will grow out.
Cancer cells often have more than the normal number of centrosomes. They also are aneuploid (have abnormal numbers of chromosomes), and considering the role of centrosomes in chromosome movement, it is tempting to think that the two phenomena are related. Mutations in the tumor suppressor gene p53 seem to predispose the cell to excess replication of the centrosomes. Chromosome movement in mitosis also involves polymerization and depolymerization of microtubules.
Because the hallmark of cancer cells is uncontrolled mitosis, both vincristine and Taxol are used as anticancer drugs. Vincristine, a drug found in the Madagascar periwinkle (a wildflower), binds to tubulin dimers preventing the assembly of microtubules. This halts cells in metaphase of mitosis and Taxol, a drug found in the bark of the Pacific yew, prevents depolymerization of the microtubules of the spindle fiber. This, in turn, stops chromosome movement, and thus prevents the completion of mitosis.