
14.3: Seed Plants - Gymnosperms
- Page ID
- 7059

The first plants to colonize land were most likely closely related to modern-day mosses (bryophytes) and are thought to have appeared about 500 million years ago. They were followed by liverworts (also bryophytes) and primitive vascular plants, the pterophytes, from which modern ferns are derived. The life cycle of bryophytes and pterophytes is characterized by the alternation of generations. The completion of the life cycle requires water, as the male gametes must swim to the female gametes. The male gametophyte releases sperm, which must swim—propelled by their flagella—to reach and fertilize the female gamete or egg. After fertilization, the zygote matures and grows into a sporophyte, which in turn will form sporangia, or "spore vessels,” in which mother cells undergo meiosis and produce haploid spores. The release of spores in a suitable environment will lead to germination and a new generation of gametophytes.
The Evolution of Seed Plants
In seed plants, the evolutionary trend led to a dominant sporophyte generation, in which the larger and more ecologically significant generation for a species is the diploid plant. At the same time, the trend led to a reduction in the size of the gametophyte, from a conspicuous structure to a microscopic cluster of cells enclosed in the tissues of the sporophyte. Lower vascular plants, such as club mosses and ferns, are mostly homosporous (produce only one type of spore). In contrast, all seed plants, or spermatophytes, are heterosporous, forming two types of spores: megaspores (female) and microspores (male). Megaspores develop into female gametophytes that produce eggs, and microspores mature into male gametophytes that generate sperm. Because the gametophytes mature within the spores, they are not free-living, as are the gametophytes of other seedless vascular plants. Heterosporous seedless plants are seen as the evolutionary forerunners of seed plants.
Seeds and pollen—two adaptations to drought—distinguish seed plants from other (seedless) vascular plants. Both adaptations were critical to the colonization of land. Fossils place the earliest distinct seed plants at about 350 million years ago. The earliest reliable record of gymnosperms dates their appearance to the Carboniferous period (359–299 million years ago). Gymnosperms were preceded by the progymnosperms (“first naked seed plants”). This was a transitional group of plants that superficially resembled conifers (“cone bearers”) because they produced wood from the secondary growth of the vascular tissues; however, they still reproduced like ferns, releasing spores to the environment. In the Mesozoic era (251–65.5 million years ago), gymnosperms dominated the landscape. Angiosperms took over by the middle of the Cretaceous period (145.5–65.5 million years ago) in the late Mesozoic era, and have since become the most abundant plant group in most terrestrial biomes.
The two innovative structures of pollen and seed allowed seed plants to break their dependence on water for reproduction and development of the embryo, and to conquer dry land. The pollen grains carry the male gametes of the plant. The small haploid (1n) cells are encased in a protective coat that prevents desiccation (drying out) and mechanical damage. Pollen can travel far from the sporophyte that bore it, spreading the plant’s genes and avoiding competition with other plants. The seed offers the embryo protection, nourishment and a mechanism to maintain dormancy for tens or even thousands of years, allowing it to survive in a harsh environment and ensuring germination when growth conditions are optimal. Seeds allow plants to disperse the next generation through both space and time. With such evolutionary advantages, seed plants have become the most successful and familiar group of plants.
Gymnosperms
Gymnosperms (“naked seed”) are a diverse group of seed plants and are paraphyletic. Paraphyletic groups do not include descendants of a single common ancestor. Gymnosperm characteristics include naked seeds, separate female and male gametes, pollination by wind, and tracheids, which transport water and solutes in the vascular system.
Life Cycle of a Conifer
Pine trees are conifers and carry both male and female sporophylls on the same plant. Like all gymnosperms, pines are heterosporous and produce male microspores and female megaspores. In the male cones, or staminate cones, the microsporocytes give rise to microspores by meiosis. The microspores then develop into pollen grains. Each pollen grain contains two cells: one generative cell that will divide into two sperm, and a second cell that will become the pollen tube cell. In the spring, pine trees release large amounts of yellow pollen, which is carried by the wind. Some gametophytes will land on a female cone. The pollen tube grows from the pollen grain slowly, and the generative cell in the pollen grain divides into two sperm cells by mitosis. One of the sperm cells will finally unite its haploid nucleus with the haploid nucleus of an egg cell in the process of fertilization.
Female cones, or ovulate cones, contain two ovules per scale. One megasporocyteundergoes meiosis in each ovule. Only a single surviving haploid cell will develop into a female multicellular gametophyte that encloses an egg. On fertilization, the zygote will give rise to the embryo, which is enclosed in a seed coat of tissue from the parent plant. Fertilization and seed development is a long process in pine trees—it may take up to two years after pollination. The seed that is formed contains three generations of tissues: the seed coat that originates from the parent plant tissue, the female gametophyte that will provide nutrients, and the embryo itself. Figure \(\PageIndex{1}\) illustrates the life cycle of a conifer.
ART CONNECTION
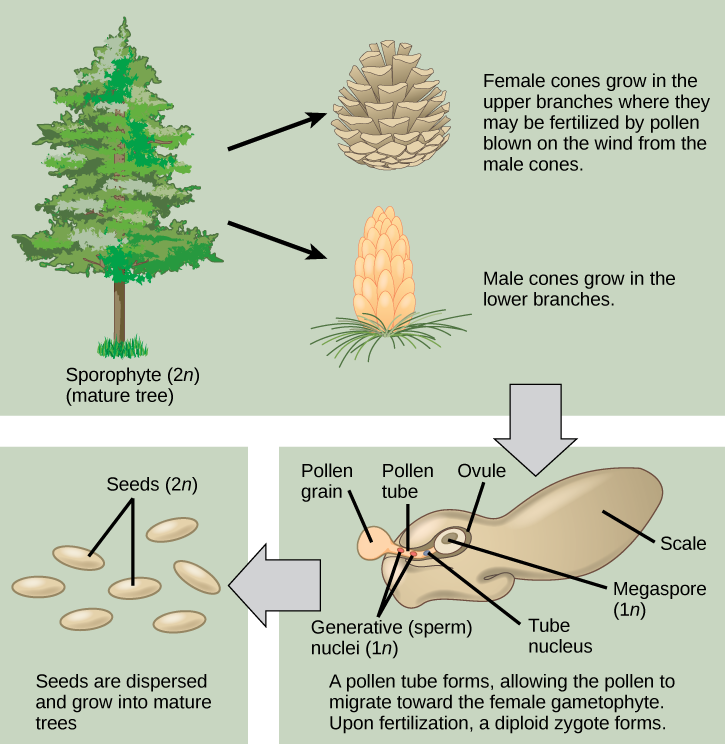
At what stage does the diploid zygote form?
- when the female cone begins to bud from the tree
- when the sperm nucleus and the egg nucleus fuse
- when the seeds drop from the tree
- when the pollen tube begins to grow
CONCEPT IN ACTION
Watch this video to see the process of seed production in gymnosperms.
Diversity of Gymnosperms
Modern gymnosperms are classified into four major divisions and comprise about 1,000 described species. Coniferophyta, Cycadophyta, and Ginkgophyta are similar in their production of secondary cambium (cells that generate the vascular system of the trunk or stem) and their pattern of seed development, but are not closely related phylogenetically to each other. Gnetophyta are considered the closest group to angiosperms because they produce true xylem tissue that contains both tracheids and vessel elements.
Conifers
Conifers are the dominant phylum of gymnosperms, with the most variety of species. Most are tall trees that usually bear scale-like or needle-like leaves. The thin shape of the needles and their waxy cuticle limits water loss through transpiration. Snow slides easily off needle-shaped leaves, keeping the load light and decreasing breaking of branches. These adaptations to cold and dry weather explain the predominance of conifers at high altitudes and in cold climates. Conifers include familiar evergreen trees, such as pines, spruces, firs, cedars, sequoias, and yews (Figure \(\PageIndex{2}\)). A few species are deciduous and lose their leaves all at once in fall. The European larch and the tamarack are examples of deciduous conifers. Many coniferous trees are harvested for paper pulp and timber. The wood of conifers is more primitive than the wood of angiosperms; it contains tracheids, but no vessel elements, and is referred to as “soft wood.”

Cycads
Cycads thrive in mild climates and are often mistaken for palms because of the shape of their large, compound leaves. They bear large cones, and unusually for gymnosperms, may be pollinated by beetles, rather than wind. They dominated the landscape during the age of dinosaurs in the Mesozoic era (251–65.5 million years ago). Only a hundred or so cycad species persisted to modern times. They face possible extinction, and several species are protected through international conventions. Because of their attractive shape, they are often used as ornamental plants in gardens (Figure \(\PageIndex{3}\)).

Gingkophytes
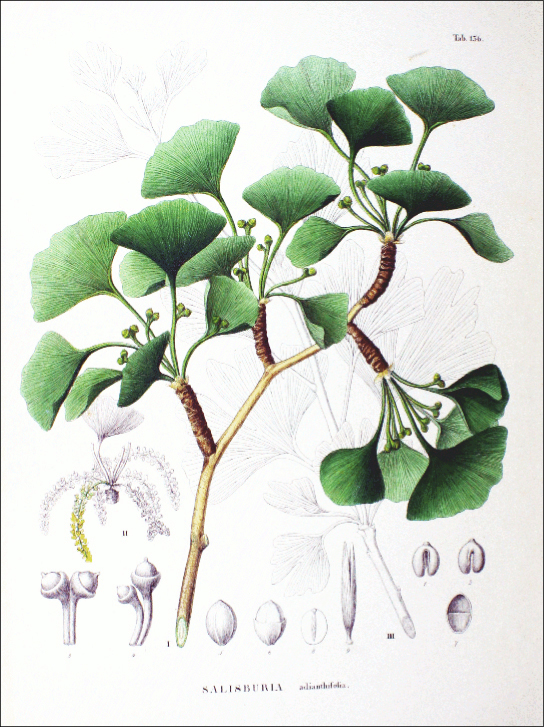
The single surviving species of ginkgophyte is the Ginkgo biloba (Figure \(\PageIndex{4}\)). Its fan-shaped leaves, unique among seed plants because they feature a dichotomous venation pattern, turn yellow in autumn and fall from the plant. For centuries, Buddhist monks cultivated Ginkgo biloba, ensuring its preservation. It is planted in public spaces because it is unusually resistant to pollution. Male and female organs are found on separate plants. Usually, only male trees are planted by gardeners because the seeds produced by the female plant have an off-putting smell of rancid butter.
Gnetophytes
Gnetophytes are the closest relatives to modern angiosperms, and include three dissimilar genera of plants. Like angiosperms, they have broad leaves. Gnetum species are mostly vines in tropical and subtropical zones. The single species of Welwitschia is an unusual, low-growing plant found in the deserts of Namibia and Angola. It may live for up to 2000 years. The genus Ephedra is represented in North America in dry areas of the southwestern United States and Mexico (Figure \(\PageIndex{5}\)). Ephedra’s small, scale-like leaves are the source of the compound ephedrine, which is used in medicine as a potent decongestant. Because ephedrine is similar to amphetamines, both in chemical structure and neurological effects, its use is restricted to prescription drugs. Like angiosperms, but unlike other gymnosperms, all gnetophytes possess vessel elements in their xylem.

CONCEPT IN ACTION
Watch this BBC video describing the amazing strangeness of Welwitschia.
Summary
Gymnosperms are heterosporous seed plants that produce naked seeds. They appeared in the Carboniferous period (359–299 million years ago) and were the dominant plant life during the Mesozoic era (251–65.5 million years ago). Modern-day gymnosperms belong to four divisions. The division Coniferophyta—the conifers—are the predominant woody plants at high altitudes and latitudes. Cycads resemble palm trees and grow in tropical climates. Gingko biloba is the only species of the division Gingkophyta. The last division, the Gnetophytes, is a diverse group of species that produce vessel elements in their wood.
Art Connections
Figure \(\PageIndex{1}\): At what stage does the diploid zygote form?
A. When the female cone begins to bud from the tree
B. When the sperm nucleus and the egg nucleus fuse
C. When the seeds drop from the tree
D. When the pollen tube begins to grow
- Answer
-
B. The diploid zygote forms after the pollen tube has finished forming so that the male generative nucleus (sperm) can fuse with the female egg.
Glossary
- cone
- the ovulate strobilus on gymnosperms that contains ovules
- conifer
- the dominant division of gymnosperms with the most variety of species
- cycad
- a division of gymnosperms that grow in tropical climates and resemble palm trees
- gingkophyte
- a division of gymnosperm with one living species, the Gingko biloba, a tree with fan-shaped leaves
- gnetophyte
- a division of gymnosperms with varied morphological features that produce vessel elements in their woody tissues
- gymnosperm
- a seed plant with naked seeds (seeds exposed on modified leaves or in cones)
- megasporocyte
- a megaspore mother cell; larger spore that germinates into a female gametophyte in a heterosporous plant
- microsporocyte
- smaller spore that produces a male gametophyte in a heterosporous plant
Contributors and Attributions
Samantha Fowler (Clayton State University), Rebecca Roush (Sandhills Community College), James Wise (Hampton University). Original content by OpenStax (CC BY 4.0; Access for free at https://cnx.org/contents/b3c1e1d2-83...4-e119a8aafbdd).