
16.4E: Photoperiodism and Phytochrome
- Page ID
- 5806

Many angiosperms flower at about the same time every year. This occurs even though they may have started growing at different times. Their flowering is a response to the changing length of day and night as the season progresses. The phenomenon is called photoperiodism. It helps promote cross pollination.
In 1920 two employees of the U. S. Department of Agriculture, W. W. Garner and H. A. Allard, discovered a mutation in tobacco - a variety called Maryland Mammoth - that prevented the plant from flowering in the summer as normal tobacco plants do. Maryland Mammoth would not bloom until late December. Experimenting with artificial lighting in winter and artificial darkening in summer, they found that Maryland Mammoth was affected by photoperiod. Because it would flower only when exposed to short periods of light, they called it a short-day plant. Some other short-day plants are
- chrysanthemums (bloom in the fall)
- rice (Oryza sativa)
- poinsettias
- morning glory (Pharbitis nil)
- the cocklebur (Xanthium)
Some plants such as spinach, Arabidopsis, sugar beet and the radish flower only after exposure to long days and hence are called long-day plants. Still other plants, e.g. the tomato, are day neutral; that is, flowering is not regulated by photoperiod.
Photoperiodism also explains why some plant species can be grown only in a certain latitude. Spinach, a long-day plant, cannot flower in the tropics because the days never get long enough (14 hours). Ragweed, a short-day plant, fails to thrive in northern Maine because by the time the days become short enough to initiate flowering, a killing frost in apt to occur before reproduction and the formation of seeds is completed.
Photoperiodism in a Short-Day Plant
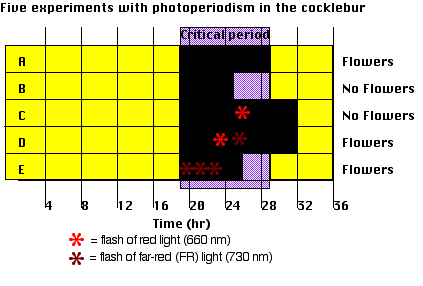
Experiments with the cocklebur have shown that the term short-day is something of a misnomer; what the cocklebur needs is a sufficiently long night.
- Cockleburs (adapted to the latitude of Michigan) will flower only if they have been kept in the dark for at least 8.5 hours — the critical period. (A and B).
- Interruption of an otherwise long night by light — red (660 nm) rays are particularly effective — prevents flowering. (C) unless
- it is followed by irradiation with far-red (730 nm) light (D).
- An intense exposure to far-red light at the start of the night reduces the dark requirement by 2 hours (E).
These response are mediated by phytochrome.
Phytochrome
- Phytochrome is a homodimer: two identical protein molecules each conjugated to a light-absorbing molecule (compare rhodopsin).
- Plants make 5 phytochromes: PhyA, PhyB, as well as C, D, and E.
- There is some redundancy in function of the different phytochromes, but there also seem to be functions that are unique to one or another. The phytochromes also differ in their absorption spectrum; that is, which wavelengths (e.g., red vs. far-red) they absorb best.
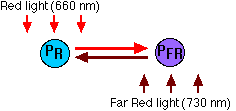
- Phytochromes exist in two interconvertible forms
- PR because it absorbs red (R; 660 nm) light
- PFR because it absorbs far-red (FR; 730 nm) light
- These are the relationships:
- Absorption of red light by PR converts it into PFR.
- Absorption of far-red light by PFR converts it into PR.
- In the dark, PFR spontaneously converts back to PR.
The Hourglass Model
The behavior of phytochrome provided the first model - called the hourglass model - of the mechanism of photoperiodism in short-day plants.
- Sunlight is richer in red (660 nm) than far-red (730 nm) light, so at sundown all the phytochrome is PFR.
- During the night, the PFR converts back to PR.
- The PR form is needed for the release of the flowering signal.
- Therefore, the cocklebur needs 8.5 hours of darkness in which to
- convert all the PFR present at sundown into PR
- carry out the supplementary reactions leading to the release of the flowering signal ("florigen").
- If this process is interrupted by a flash of 660-nm light, the PR is immediately reconverted to PFR and the night's work is undone (C)
- A subsequent exposure to far-red (730 nm) light converts the pigment back to PR and the steps leading to the release of "florigen" can be completed (D)
- Exposure to intense far-red light at the beginning of the night sets the clock ahead about 2 hours or so by eliminating the need for the spontaneous conversion of PFR to PR (E).
The Circadian Rhythm Model
Recent work mostly in the long-day plant, Arabidopsis - supports a different model of photoperiodism. This work suggests that the photoperiodic response is governed by the interaction of daylight with innate circadian rhythms of the plant.
- Virtually all eukaryotes have innate circadian rhythms.
- These are rhythms of biological activities that fluctuate over a period of approximately 24 hours (L. circa = about; dies = day) even under constant environmental conditions (e.g. continuous darkness). Under constant conditions, the cycles may drift out of phase with the environment.
- However, when exposed to the environment (e.g., alternating day and night), the rhythms become entrained; that is, they now cycle in lockstep with the cycle of day and night with a period of exactly 24 hours.
- In Arabidopsis, the entrainment of the rhythms requires that light is detected by the
- phytochromes (absorb red light)
- cryptochromes (absorb blue light)
Long-Day Plants
Arabidopsis is a long-day plant and has provided many clues about the mechanism involved in this photoperiodic response. Any response to photoperiod requires a method of keeping time; that is, a clock. Plants, like so many other organisms, have an innate circadian rhythm that regulates the expression of many genes. Among these in Arabidopsis is CONSTANS (CO), a gene that encodes a zinc-finger transcription factor whose levels of mRNA rise and fall with a circadian rhythm.
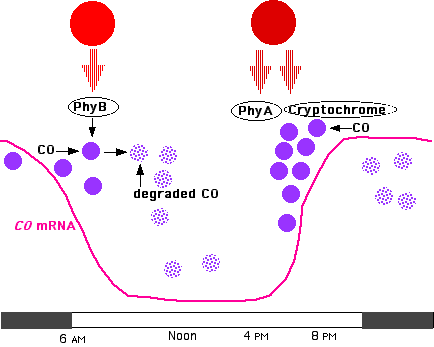
Translation of CONSTANS mRNA produces the transcription factor that turns on a number of genes, including FLOWERING LOCUS T (FT), a gene needed to start the conversion of apical buds into flower buds.
CONSTANS messenger RNA (mRNA) is abundant early in the morning, declines during the middle part of the day and then rises to another peak late in the afternoon.
However,
- the CONSTANS protein is quickly degraded (in proteasomes) during the morning and middle part of the day and also during the night.
- The degradation triggered by morning light (rich in 660 nm rays) is mediated by phytochrome B (PhyB).
- By late in the afternoon, if the day has been long enough,
- transcription of the CO gene increases producing a rise in CO mRNA
- translation of the CO mRNA produces more CO protein which is
- no longer degraded.
- These effects are mediated by the absorption of
- red (enriched in far-red) light by phytochrome A (PhyA)
- blue light by cryptochrome.
Now with the CONSTANS protein accumulating, it is available to turn on the gene transcription (e.g., FT) needed for the induction of the flowering. In short days, with darkness falling before the rise in CONSTANS mRNA, there is not enough CONSTANS protein synthesized to induce flowering. So flowering in Arabidopsis seems to require the interaction of daylight perceived by phytochromes and cryptochromes with the intrinsic circadian rhythm of CONSTANS expression.
Short-Day Plants
The roles of circadian rhythms and light in short-day plants are not yet as well understood. Studies with rice, a short-day plant, suggests that the mechanism described for Arabidopsis may work there as well but with CONSTANS acting as a suppressor of FLOWERING LOCUS T and thus as an inhibitor of flowering under long days.
Trees
Photoperiodism not only controls flowering in some trees but also stops vegetative growth and promotes the setting of winter buds as the days grow shorter in the autumn. In aspens (Populus sp.) this control is mediated by CONSTANS and FLOWERING LOCUS T.
Contributors and Attributions
John W. Kimball. This content is distributed under a Creative Commons Attribution 3.0 Unported (CC BY 3.0) license and made possible by funding from The Saylor Foundation.