
15.4K: Organ Transplants
- Page ID
- 5447

New parts for old
Many organs and tissues are now routinely transplanted from one human to another. Except for the rare cases where the donor and recipient are monozygotic ("identical") twins, such grafts are called allografts.
- kidney
- Living donors can be used because they have two kidneys and can get along with only one.
- heart
- For patients with failing hearts (often because of inherited defects). Only cadavers can be used as donors.
- lungs
- Usually transplanted along with a heart. Some attempts have been made with portions of lungs from living donors.
- pancreas
- For people with Type 1 diabetes mellitus.
- liver
- For irreversible liver failure (e.g., from toxins, hepatitis B infection).
- skin
- For burns; usually taken from elsewhere on the patient's own body.
- cornea
- To restore sight; taken from cadavers.
- blood
- To temporarily restore blood volume.
- bone marrow
- As a source of blood ("hematopoietic") stem cells to repopulate the patient's own marrow that is
- congenitally deficient in its ability to make one or more kinds of blood cells — Example: severe combined immunodeficiency (SCID)
- has been destroyed by cancer therapy.
- As a source of blood ("hematopoietic") stem cells to repopulate the patient's own marrow that is
- cord blood
- Blood drained (through the umbilical cord) from the placenta of newborn infants. A convenient source of blood stem cells.
- ovary
- Has restored fertility and produced healthy babies but so far only when donor and recipient were monozygotic (identical) twins.
The Problems
- Graft rejection
- The patient's immune system "sees" an allograft as foreign (antigenic) and mounts an immune response against it.
- Graft-versus-host disease (GVHD)
- T cells in the graft "see" the tissues of the recipient as foreign antigens and mount an immune attack against them. This a particularly serious problem with grafts of bone marrow because of the many T cells in it.
- Infections
- Attempts to suppress the immune response to avoid graft rejection and GVHD weaken the ability of the body to combat infectious agents (bacteria, viruses, fungi).
- More rarely, the donated organ may be infected and transmit the agent to the recipient. Tuberculosis, rabies, syphilis, hepatitis B, HIV-1, and several other diseases have been transmitted in this way. Potential organ donors are now routinely tested for evidence of infection by HIV-1 and -2, HTLV-1 and -2, hepatitis B and C (HBV, HCV), human cytomegalovirus (HCMV) and Epstein-Barr virus (EBV) as well as by Treponema pallidum (syphilis).
- Cancer
- Suppressing the host's immune responses also increases the risk of cancer.
Coping with the Immunological Problems
- Use the patient's own tissue when possible (skin, bone marrow, blood vessels).
- Use tissue from an "identical" (monozygotic) twin in the very rare cases that one is available. Being genetically identical, the recipient sees the transplant as "self", not as foreign, and does not mount an attack against it. The first successful kidney transplants (done in the mid 1950s) were between identical twins, and both donors and recipients went on to lead normal lives.
- Use an "immunologically privileged" site. These are parts of the body where the immune system is prevented from mounting an attack. They include the eye, testes, and brain, but only the eye's privileged status has so far been exploited (for corneal grafts).
- Use a relative, preferably a sibling, as the donor. While never identical, they may have inherited some of the same histocompatibility antigens so the recipient's immune response may not be as strong as it otherwise would be.
- Tissue-typing. Determine the histocompatibility antigens of both recipient and potential donor and use the organ with the fewest mismatches.
- Immunosuppression. Use immunosuppressive agents to blunt the recipient's immune response. Invariably required for all allografts.
Tissue Typing
The strongest antigens expressed by tissues are the class I and class II histocompatibility molecules. These are encoded by an array of genes on chromosome 6 called the major histocompatibility complex (MHC).
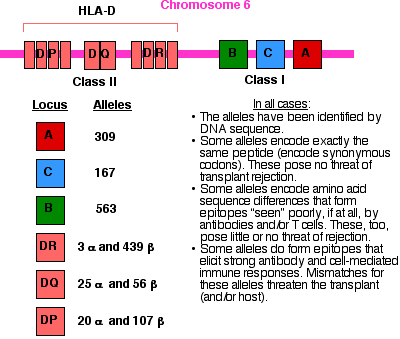
Class I molecules consist of a transmembrane protein to which are attached (noncovalently), a molecule of beta-2 microglobulin, and a short peptide. The class I transmembrane proteins are encoded by three loci: HLA-A, HLA-B, and HLA-C. Class I molecules are expressed at the surface of almost all the cells of the body (except for red blood cells and the cells of the central nervous system).
Class II molecules consist of two transmembrane polypeptides: an alpha (α) chain and beta (β) chain between which is nestled (noncovalently) a short peptide. The alpha and beta chains are encoded by clusters of loci in the region of chromosome 6 designated HLA-D. Unlike class I molecules, class II molecules are expressed on only a few types of cells, chiefly antigen-presenting cells (APCs) such as dendritic cells and macrophages, as well as other cells where inflammation is occurring.
Why so many MHC alleles
The genes of the MHC are the most polymorphic known. The graphic above shows the latest counts of alleles found at each locus in the human population. Of course, any one human can inherit a maximum of two alleles at each locus. The diversity of alleles in the population makes possible thousands of different combinations. In a study of 1000 blood and organ donors in San Francisco that were typed for HLA-A and HLA-B,
- Over half the group had a combination that was unique.
- Another 111 donors had a set of these molecules that they shared with only one other person in the group.
- The most frequent phenotype (HLA-A1, HLA-A3, HLA-B7, and HLA-B8) was found in 11 donors.
The extreme polymorphism of the MHC did not evolve to frustrate transplant surgeons and their patients.
Why, then?
The function of class I and class II molecules is to "present" antigenic peptides to the T cells of the immune system. The peptides — usually about 9 amino acids long — are bound by noncovalent forces in a groove at the surface of the MHC molecule. These peptides can include fragments of protein antigens derived from intracellular pathogens (e.g. viruses). Different pathogens generate different antigenic fragments. Different MHC molecules differ in the efficiency with which they bind particular sequences of amino acids in these fragments. Therefore, we might expect that some MHC products would be better than others at presenting pathogen antigens to the immune system.
One piece of evidence: People infected with the human immunodeficiency virus (HIV-1) who have one particular HLA-B molecule are more resistant to developing a full-blown case of AIDS than those with other HLA-B molecules (even though some of these differ by only a single amino acid).
Another piece of evidence: Wegner and colleagues reported in the 5 September 2003 issue of Science the results of a direct test of this hypothesis. They exposed groups of sticklebacks — differing in the number of their class II alleles — to three types of parasite (all at once). They used two species of parasitic nematode and one species of trematode). Those fish with 5–6 different alleles resisted infection better than those with fewer (or more).
So it may well be that the great diversity of class I and class II alleles in the human population has helped ensure that no single pathogen can wipe out the entire population.
Techniques of tissue typing
Most tissue typing is done using serological methods: antibodies specific for those HLA antigens that have been identified in the human population. A reaction between cells of the subject and, for example, anti-HLA-A28 antibodies and HLA-A9 antibodies — but no other antibodies — establishes the phenotype. At the present time, routine typing is limited to establishing the phenotype at HLA-A, HLA-B, and HLA-DR.
Coming into wider use is DNA typing, especially for HLA-D antigens. It promises to improve the sensitivity and specificity of tissue typing. The totals of numbers of alleles at each HLA locus given in the graphic above are based on DNA typing.
How useful is tissue typing?
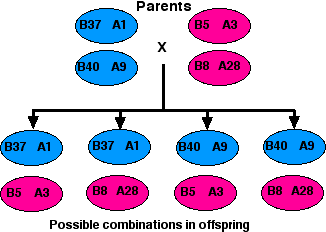
So what hope do these data hold for the dialysis patient awaiting a kidney transplant? If the patient has a large family of willing donors, the odds for a good match are not bad because of the tight linkage of the HLA loci.
Assuming that no crossing over occurs within the HLA region of either the mother's or the father's two number 6 chromosomes, there are four possible combinations in which they may transmit their alleles to their children. So even if the parents carry different alleles at each locus (which is often the case), there is a 1:4 probability that any one of their children will be an exact match with any other. (Only the HLA-A and HLA-B antigens are shown here, but the tight linkage of the entire HLA region makes it likely that all the loci on each chromosome will be passed on as a block.)
But most organs are transplanted from cadavers to complete strangers. In the United States, the program is monitored by the United Network for Organ Sharing (UNOS). Tissue typing is usually limited to looking for 6 HLA antigens: the two each at HLA-A, HLA-B, and HLA-DR. If only one antigen is found at a locus, it means that either the tissue is homozygous for that allele or no reagent exists yet to detect the second allele. This table shows the results of one study of several thousand kidney recipients.
| Number of HLA mismatches |
% kidneys surviving after 5 years |
|---|---|
| 0 | 68 |
| 1 | 61 |
| 2 | 61 |
| 3 | 58 |
| 4 | 58 |
| 5 | 57 |
| 6 | 56 |
The results tell us that:
- Having no mismatches provides a clear, but modest, advantage over mismatched kidneys. (This advantage is cumulative: at 17 years, 50% of the kidneys with no mismatches are still functioning while 50% of those with one or more mismatches have been lost after 8 years.)
- However, the incremental disadvantage of additional mismatches is small. In fact, the procedures to prevent rejection are now sufficiently good that ~90% of all kidneys — even those with all loci mismatched — can be expected to be functioning at the end of the first year.
Minor histocompatibility antigens
Even if it were possible to match donor and recipient at every locus of the MHC, some tissue incompatibility would still remain (except between identical twins). Few of the antigens responsible have been identified, but they include:
- H-Y, an antigen encoded on the Y chromosome and thus present in male, but not female, tissue
- HA-2, an antigen derived from the contractile protein myosin.
The number and variety of histocompatibility antigens tell us that probably no two humans (again, except for identical twins) exist on earth with perfectly compatible tissues. Therefore successful transplantation of allografts requires some degree of immunosuppression to avoid graft rejection.
Graft-versus-host disease (GVHD)
Allografts that contain T cells of the donor can cause graft-versus-host disease (GVHD). The T cells in the transplant see the host as "foreign" and proceed to mount a widespread attack against the tissues of the host. GVHD is an especially serious problem with grafts of bone marrow (the source of all blood cells) and cord blood. Even when the donor and recipient have identical HLA alleles, grafts of bone marrow often cause GVHD because of differences in their minor histocompatibility antigens.
Some cancer patients are now deliberately treated so vigorously with radiation and chemotherapy that their bone marrow is destroyed along with their cancer cells. In order to survive, these patients must be given stem cells to repopulate their marrow after their therapy. In some cases, their own bone marrow is used. Some of it is removed prior to the onset of treatment of the patient and is itself treated to remove any cancer cells that may be lurking in it. If allografted bone marrow is required, there is a strong danger of GVHD. If the GVHD can be controlled, the stem cells should eventually establish themselves in the bone marrow of their new host and in due course generate some or even all of the patient's blood cells. Cord blood — another source of stem cells — presents less of a risk of serious GVHD even from a donor with HLA molecules not present in the recipient. This is because cord blood does not contain any mature T cells.
In mice, GVHD can be minimized by injecting large numbers of regulatory T cells, but for humans, control of GVHD — like control of graft rejection — still depends on the use of immunosuppression.
Immunosuppression
Immunosuppression is the treatment of the patient with agents that inhibit the immune response. The following is the list of immunosuppressants currently used.
Purine analogs
These are relatives of the purines used in DNA synthesis. Because they interfere with DNA synthesis, they interfere with the rapid cell proliferation needed for immune responses. Azathioprine (trade name = Imuran) is a widely-used purine analog.
Unfortunately, these drugs also interfere with the many other tissues that depend on rapid cell division (e.g., lining of the intestine, hair follicles) so they have many unpleasant side effects. Therefore, the search for agents that specifically target immune cells goes on.
Corticosteroids
These relatives of cortisol interfere with a transcription factor needed to turn on the genes for T cells to become activated. Prednisone and prednisolone are most commonly used.
Tacrolimus (Prograf®) and cyclosporine (Neoral®)
These are natural products isolated from microbial cultures. They inhibit the signaling pathway used by T cells to turn on their genes for activation, e.g., for IL-2 secretion.
Rapamycin
This is a macrolide antibiotic produced by an actinomycete found on Easter Island (which the inhabitants call Rapa Nui — hence the name). Rapamycin inhibits T cell proliferation, and shows great promise in reducing the problems of transplant rejection.
Rapamycin is also known as sirolimus and is sold under the trade name Rapamune.
Mycophenolate mofetil
This small molecule inhibits an enzyme needed by B and T cells for purine synthesis. Other types of cells are not dependent on the enzyme so side effects are mild. The trade name is CellCept.
Antithymocyte globulin (ATG)
This preparation contain antibodies — raised in horses or rabbits — directed against T cells.
Monoclonal antibodies
Several preparations are now used:
- Muromonab-CD3 (OKT3) and two humanized anti-CD3 monoclonals. They bind to the CD3 molecule on the surface of T cells.
- Daclizumab and basiliximab. Target the IL-2 receptor and thus inhibit only activated T cells.
- Alemtuzumab (Campath-1H®). Binds to CD52, a molecule found on lymphocytes and depletes both T cells and B cells.
Belatacept
This is a protein, produced by recombinant DNA technology, that combines
- the extracellular portion of CTLA-4 ("cytotoxic T-lymphocyte-associated antigen 4", one of the ligands for B7) with
- the Fc region (the C-terminal two-thirds of the constant region) of a human IgG1 antibody.
It blocks the "Signal Two" needed to activate T cells.
Side effects of immunosuppression
They are serious.
Infections
The immune system is vital to protect us against infectious agents (bacteria, viruses, fungi). So infection is a frequent side effect of immunosuppression in transplant recipients. Fortunately, the infections can usually be controlled by the appropriate antibiotic, antiviral drug, etc.
Cancer
5% or more of transplant recipients will develop cancer within a few years of receiving their allograft. This may not seem to represent a great risk, but it does represent a 100-fold increase in risk compared to the general population. Allograft recipients are particularly prone to developing skin cancers and lymphomas. Curiously, transplant recipients do not seem to be at any greater risk for developing the most common types of cancer in the rest of the population: cancers of the lung, breast, colon, and prostate. One exception: recipients of allografted bone marrow run a slightly, but significantly, higher (2–3 fold) risk of developing these types of tumors. In most cases, these cancers arise from a cell of the host. But in some cases of melanoma and Kaposi's sarcoma the cancer cells were present in the graft and proliferated in the immunosuppressed host.
Things that can be done to help in such cases include stopping the immunosuppression. The chief culprit seems to be the immunosuppression that these patients have been receiving. In most cases,this leads to regression of the cancer, but often rejection of the transplant as well. The choice is usually clear for patients with allografted kidneys; they can go back on dialysis and anticipate receiving another kidney at a future date. But what of the cancer patient with a heart transplant?
Future prospects for transplantation
Although organ transplants have helped thousands of people, much remains to be done. In particular, ways need to be found to
- increase the number of available organs (the need now far exceeds the supply)
- find more precise methods of immunosuppression in order to prevent rejection without the dangerous side effects of infection and cancer.
Both these problems may be helped by xenotransplantation.
Xenotransplantation
Xenotransplantation is the use of organs from other animals. A number of attempts have been made to use hearts, livers, and kidneys from such primates as chimpanzees and baboons — so far with limited success. One reason is that xenotransplants usually are attacked immediately by antibodies of the host resulting in hyperacute rejection. But perhaps the use of pigs as organ donors will be feasible.
- Their organs are about the right size for use in humans.
- They can be made transgenic for molecules that may circumvent
- hyperacute rejection (by knocking out the genes responsible for cell-surface antigens that humans have preformed antibodies against);
- the chronic, T-cell-mediated, rejection that plagues all allografts.
- They can be produced in the numbers needed.
However, pigs contain retroviruses (called PERV = porcine endogenous retrovirus), and there is fear that these might infect the human recipient.
Only a few transplants of pig tissue into humans have been done to date: skin grafts and grafts of pancreatic islets. A larger number of people have been temporarily hooked up to pig organs or "bioreactors" containing pig cells to provide support for their failing spleen, liver, or kidneys. Most of these recipients have been monitored for signs of infection by PERV and — even though PERV can infect human cells growing in culture — there is no evidence that any of these people exposed to pig tissue have become infected.
Is xenotransplantation safe?
Pigs are not the only animals that contain latent viruses in their cells. Could the viruses in the cells of other kinds of animal donors infect the transplant recipient? start an epidemic? The danger seems greater for xenotransplants from other primates. (HIV, the retrovirus that causes AIDS appears to have entered humans from a primate host)
For these reasons, many biologists are urging that transplant surgeons proceed cautiously with xenotransplants.
Immune Privilege
It has long been know that certain sites in the body, for example,
- the anterior chamber of the eye
- the testes
- the brain
are "privileged". They are protected from attack by the immune system.
This can cause problems. Several cases have emerged of survivors of Ebola who no longer have Ebola virus in their blood and are symptom-free but still retain live virus in such privileged sites as brain, testis, and aqueous humor of the eye where the virus has escaped attack by the immune system.
Many factors are involved in immune privilege, such as
- tight junctions between the cells of the tissue
- little expression of class I histocompatibility molecules
- expression of the Fas ligand, FasL.
The presence of FasL on their surface protects cells in privileged locations from immune attack because when threatened by a cytotoxic T cell, they force the T cell to commit suicide by apoptosis. Activated cytotoxic T cells express Fas on their surface. When they engage (with their T cell receptor) a privileged cell expressing FasL, instead of killing the target, the target kills them!
So if the organs of transgenic pigs can be made to expresses human FasL, perhaps they will be resistant to T-cell mediated rejection.
The human placenta also enjoys immune privilege. It is almost as foreign to the mother as a kidney transplant from her husband would be, but unlike the kidney, she will not reject it (at least not for 9 months). In lab rats, the embryos (and the mother's endometrium) secrete corticotropin-releasing hormone (CRH). This hormone induces the expression of Fas ligand (FasL) on the cells of the placenta. Activated T cells express Fas, so any threatening T cells would commit suicide by apoptosis when they encounter FasL on their target.