
5.5: DNA Replication
- Page ID
- 4743

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \) \( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)\(\newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\) \( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\) \( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\) \( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\) \( \newcommand{\Span}{\mathrm{span}}\) \(\newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\) \( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\) \( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\) \( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\) \( \newcommand{\Span}{\mathrm{span}}\)\(\newcommand{\AA}{\unicode[.8,0]{x212B}}\)
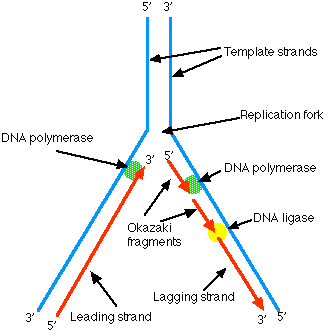
With their multiple origins, how does the eukaryotic cell know which origins have been already replicated and which still await replication?
An observation: When a cell in G2 of the cell cycle is fused with a cell in S phase, the DNA of the G2 nucleus does not begin replicating again even though replication is proceeding normally in the S-phase nucleus. Not until mitosis is completed, can freshly-synthesized DNA be replicated again.
Two control mechanisms have been identified — one positive and one negative. This redundancy probably reflects the crucial importance of precise replication to the integrity of the genome.
Licensing: positive control of replication
In order to be replicated, each origin of replication must be bound by:
- an Origin Recognition Complex of proteins (ORC). These remain on the DNA throughout the process.
- Accessory proteins called licensing factors. These accumulate in the nucleus during G1 of the cell cycle. They include:
- Cdc-6 and Cdt-1, which bind to the ORC and are essential for coating the DNA with
- MCM proteins. Only DNA coated with MCM proteins (there are 6 of them) can be replicated.
Once replication begins in S phase,
- Cdc-6 and Cdt-1 leave the ORCs (the latter by ubiquination and destruction in proteasomes).
- The MCM proteins leave in front of the advancing replication fork.