
8.8: Carbohydrate Storage and Breakdown
- Page ID
- 66031

Carbohydrates are important cellular energy sources. They provide energy quickly through glycolysis and passing of intermediates to pathways, such as the citric acid cycle, amino acid metabolism (indirectly), and the pentose phosphate pathway. It is important, therefore, to understand how these important molecules are made.
Plants are notable in storing glucose for energy in the form of amylose and amylopectin (see and for structural integrity in the form of cellulose. These structures differ in that cellulose contains glucoses solely joined by beta-1,4 bonds, whereas amylose has only alpha1,4 bonds and amylopectin has alpha 1,4 and alpha 1,6 bonds.Animals store glucose primary in liver and muscle in the form of a compound related to amylopectin known as glycogen. The structural differences between glycogen and amylopectin are solely due to the frequency of the alpha 1,6 branches of glucoses. In glycogen they occur about every 10 residues instead of every 30-50, as in amylopectin.
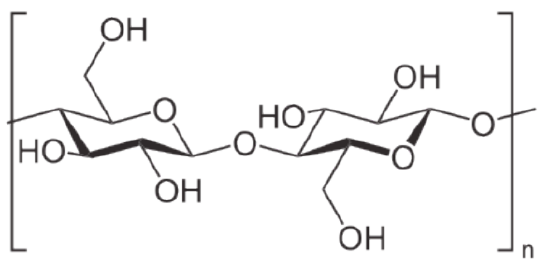
Glycogen provides an additional source of glucose besides that produced via gluconeogenesis. Because glycogen contains so many glucoses, it acts like a battery backup for the body, providing a quick source of glucose when needed and providing a place to store excess glucose when glucose concentrations in the blood rise. The branching of glycogen is an important feature of the molecule metabolically as well. Since glycogen is broken down from the "ends" of the molecule, more branches translate to more ends, and more glucose that can be released at once. Breakdown of glycogen involves
- release of glucose-1- phosphate (G1P),
- rearranging the remaining glycogen (as necessary) to permit continued breakdown, and
- conversion of G1P to G6P for further metabolism. G6P can be 1) broken down in glycolysis, 2) converted to glucose by gluconeogenesis, and 3) oxidized in the pentose phosphate pathway.
Just as in gluconeogenesis, the cell has a separate mechanism for glycogen synthesis that is distinct from glycogen breakdown. As noted previously, this allows the cell to separately control the reactions, avoiding futile cycles, and enabling a process to occur efficiently (synthesis of glycogen) that would not occur if it were simply the reversal of glycogen breakdown.

Synthesis of glycogen starts with G1P, which is converted to an 'activated' intermediate, UDP-glucose. This activated intermediate is what 'adds' the glucose to the growing glycogen chain in a reaction catalyzed by the enzyme known as glycogen synthase. Once the glucose is added to glycogen, the glycogen molecule may need to have branches inserted in it by the enzyme known as branching enzyme.
Glycogen Breakdown
Glycogen phosphorylase (sometimes simply called phosphorylase) catalyzes breakdown of glycogen into Glucose-1-Phosphate (G1P). The reaction, (see above right) that produces G1P from glycogen is a phosphorolysis, not a hydrolysis reaction. The distinction is that hydrolysis reactions use water to cleave bigger molecules into smaller ones, but phosphorolysis reactions use phosphate instead for the same purpose. Note that the phosphate is just that - it does NOT come from ATP. Since ATP is not used to put phosphate on G1P, the reaction saves the cell energy.

Glycogen phosphorylase will only act on non-reducing ends of a glycogen chain that are at least 5 glucoses away from a branch point. A second enzyme, Glycogen Debranching Enzyme (GDE), is therefore needed to convert alpha(1-6) branches to alpha(1-4) branches. GDE acts on glycogen branches that have reached their limit of hydrolysis with glycogen phosphorylase. GDE acts to transfer a trisaccharide from a 1,6 branch onto an adjacent 1,4 branch, leaving a single glucose at the 1,6 branch. Note that the enzyme also catalyzes the hydrolysis of the remaining glucose at the 1,6 branch point. Thus, the breakdown products from glycogen are G1P and glucose (mostly G1P, however). Glucose can, of course, be converted to Glucose-6-Phosphate (G6P) as the first step in glycolysis by either hexokinase or glucokinase. G1P can be converted to G6P by action of an enzyme called phosphoglucomutase. This reaction is readily reversible, allowing G6P and G1P to be interconverted as the concentration of one or the other increases. This is important, because phosphoglucomutase is needed to form G1P for glycogen biosynthesis.
Regulation of Glycogen Metabolism
Regulation of glycogen metabolism is complex, occurring both allosterically and via hormone-receptor controlled events that result in protein phosphorylation or dephosphorylation. In order to avoid a futile cycle of glycogen synthesis and breakdown simultaneously, cells have evolved an elaborate set of controls that ensure only one pathway is primarily active at a time.

Regulation of glycogen metabolism is managed by the enzymes glycogen phosphorylase and glycogen synthase. Glycogen phosphorylase is regulated by both allosteric factors (ATP, G6P, AMP, and glucose) and by covalent modification (phosphorylation/dephosphorylation). Its regulation is consistent with the energy needs of the cell. High energy substrates (ATP, G6P, glucose) allosterically inhibit GP, while low energy substrates (AMP, others) allosterically activate it.
GPa/GPb Allosteric Regulation
Glycogen phosphorylase exists in two different covalent forms – one form with phosphate (called GPa here) and one form lacking phosphate (GPb here). GPb is converted to GPa by phosphorylation by an enzyme known as phosphorylase kinase. GPa and GPb can each exist in an 'R' state and a 'T' state. For both GPa and GPb, the R state is the more active form of the enzyme. GPa's negative allosteric effector (glucose) is usually not abundant in cells, so GPa does not .ip into the T state often. There is no positive allosteric effector of GPa, so when glucose is absent, GPa automatically flips into the R (more active) state.
GPb can convert from the T state to the GPb R state by binding AMP. Unless a cell is low in energy, AMP concentration is low. Thus GPb is not converted to the R state very often. On the other hand, ATP and/or G6P are usually present at high enough concentration in cells that GPb is readily flipped into the T state.
GPa/GPb Covalent Conversion
Because the relative amounts of GPa and GPb largely govern the overall process of glycogen breakdown, it is important to understand the controls on the enzymes that interconvert GPa and GPb. This is accomplished by the enzyme Phosphorylase Kinase, which transfers phosphates from 2 ATPs to GPb to form GPa. Phosphorylase kinase has two covalent forms – phosphorylated (active) and dephosphorylated (inactive). It is phosphorylated by the enzyme Protein Kinase A (PKA). Another way to activate the enzyme is with calcium. Phosphorylase kinase is dephosphorylated by the same enzyme, phosphoprotein phosphatase, that removes phosphate from GPa.
PKA is activated by cAMP, which is, in turn produced by adenylate cyclase after activation by a G-protein. G-proteins are activated ultimately by binding of ligands to specific 7-TM receptors, also known as G-protein coupled receptors. These are discussed in greater detail in Chapter 8. Common ligands for these receptors include epinephrine (binds beta-adrenergic receptor) and glucagon (binds glucagon receptor). Epinephrine exerts it greatest effects on muscle and glucagon works preferentially on the liver.
Turning Off Glycogen Breakdown
Turning OFF signals is as important, if not more so, than turning them ON. The steps in the glycogen breakdown regulatory pathway can be reversed at several levels. First, the ligand can leave the receptor. Second, the G-proteins have an inherent GTPase activity that serves to turn them off over time. Third, cells have phosphodiesterase (inhibited by caffeine) for breaking down cAMP. Fourth, an enzyme known as phosphoprotein phosphatase can remove phosphates from phosphorylase kinase (inactivating it) AND from GPa, converting it to the much less active GPb.
Glycogen Synthesis
The anabolic pathway contrasting with glycogen breakdown is that of glycogen synthesis. Just as cells reciprocally regulate glycolysis and gluconeogenesis to prevent a futile cycle, so too do cells use reciprocal schemes to regulate glycogen breakdown and synthesis. Let us first consider the steps in glycogen synthesis. 1) Glycogen synthesis from glucose involves phosphorylation to form G6P, and isomerization to form G1P (using phosphoglucomutase common to glycogen breakdown). G1P is reacted with UTP to form UDP-glucose in a reaction catalyzed by UDP-glucose pyrophosphorylase. Glycogen synthase catalyzes synthesis of glycogen by joining carbon #1 of the UDPG-derived glucose onto the carbon #4 of the non-reducing end of a glycogen chain. to form the familiar alpha(1,4) glycogen links. Another product of the reaction is UDP.
It is also worth noting in passing that glycogen synthase will only add glucose units from UDPG onto a preexisting glycogen chain that has at least four glucose residues. Linkage of the first few glucose units to form the minimal "primer" needed for glycogen synthase recognition is catalyzed by a protein called glycogenin, which attaches to the first glucose and catalyzes linkage of the first eight glucoses by alpha(1,4) bonds. 3) The characteristic alpha(1,6) branches of glycogen are the products of an enzyme known as Branching Enzyme. Branching Enzyme breaks alpha(1,4) chains and carries the broken chain to the carbon #6 and forms an alpha(1,6) linkage.
Regulation of Glycogen Synthesis
The regulation of glycogen biosynthesis is reciprocal to that of glycogen breakdown. It also has a cascading covalent modification system similar to the glycogen breakdown system described above. In fact, part of the system is identical to glycogen breakdown. Epinephrine or glucagon signaling can stimulate adenylate cyclase to make cAMP, which activates PKA, which activates phosphorylase kinase.
In glycogen breakdown, phosphorylase kinase phosphorylates GPb to the more active form, GPa. In glycogen synthesis, protein kinase A phosphorylates the active form of glycogen synthase (GSa), and converts it into the usually inactive b form (called GSb). Note the conventions for glycogen synthase and glycogen phosphorylase. For both enzymes, the more active forms are called the 'a' forms (GPa and GSa) and the less active forms are called the 'b' forms (GPb and GSb). The major difference, however, is that GPa has a phosphate, but GSa does not and GPb has no phosphate, but GSb does. Thus phosphorylation and dephosphorylation have opposite effects on the enzymes of glycogen metabolism. This is the hallmark of reciprocal regulation. It is of note that the less active glycogen synthase form, GSb, can be activated by G6P. Recall that G6P had the exactly opposite effect on GPb.

Glycogen synthase, glycogen phosphorylase (and phosphorylase kinase) can be dephosphorylated by several enzymes called phosphatases. One of these is called Protein Phosphatase and it is activated when insulin binds to a receptor in the cell membrane. It causes PP to be activated, stimulating dephosphorylation, and thus activating glycogen synthesis and inhibiting glycogen breakdown. Again, there is reciprocal regulation of glycogen synthesis and degradation.

Maintaining Blood Glucose Levels
After a meal, blood glucose levels rise and insulin is released. It simultaneously stimulates uptake of glucose by cells and incorporation of it into glycogen by activation of glycogen synthase and inactivation of glycogen phosphorylase. When blood glucose levels fall, GPa gets activated (stimulating glycogen breakdown to raise blood glucose) and GSb is formed (stopping glycogen synthesis).
Contributors and Attributions
Dr. Kevin Ahern and Dr. Indira Rajagopal (Oregon State University)