
2020_Winter_Bis2a_Facciotti_Lecture_25
- Page ID
- 27789
Learning Objectives Associated with 2020_Winter_Bis2a_Facciotti_Lecture_25
|
Mutations

Errors occurring during DNA replication are not the only way by which mutations can arise in DNA. Mutations, variations in the nucleotide sequence of a genome, can also occur because of physical damage to DNA. Such mutations may be of two types: induced or spontaneous. Induced mutations are those that result from an exposure to chemicals, UV rays, x-rays, or some other environmental agent. Spontaneous mutations occur with no exposure to any environmental agent; they result from spontaneous biochemical reactions taking place within the cell.
Mutations may have a wide range of effects.
As we will visit later, when a mutation occurs in a protein coding region, it may have several effects. Transition or transversion mutants may lead to no change in the protein sequence (known as silent mutations), change the amino acid sequence (known as missense mutations), or create
Note: Vocabulary Watch
Note that the preceding paragraph had a lot of potentially new vocabulary - it would be a good idea to learn these terms.
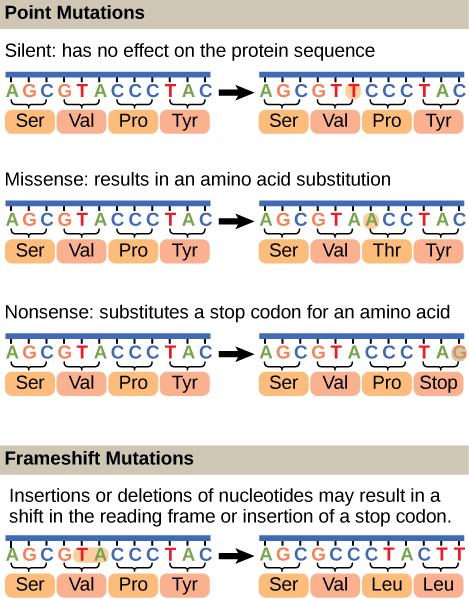
Figure 1. Mutations can lead to changes in the protein sequence encoded by the DNA.
Mutations: Some nomenclature and considerations
Mutation
Etymologically, the term mutation means a change or alteration. In genetics, a mutation is a change in the genetic material - DNA sequence - of an organism. By extension, a mutant is the organism in which a mutation has occurred. But what is the change compared to? The answer to this question is that it depends. We can make the comparison with the direct progenitor (cell or organism) or
Wild Type vs Mutant
What do we mean by "wild type"? Since the definition can depend on context, this concept is not entirely straightforward. Here are a few examples of definitions you may run into:
Possible meanings of "wild-type"
- An organism having an appearance characteristic of the species in a natural breeding population (i.e. a cheetah's spots and tear-like dark streaks that extend from the eyes to the mouth).
- The form or forms of a gene most commonly occurring in nature in a
given species. - A phenotype, genotype, or gene that predominates in a natural population of organisms or strain of organisms in contrast to that of natural or laboratory mutant forms.
- The normal, as opposed to the mutant gene or allele.
In this class we take what is common about those varying definitions and adopt the idea that "wild type" is
Possible NB Discussion  Point
Point
If you were assigning wild type traits to describe a dog, what would they be? What is the difference between a mutant trait and variation of a trait in a population of dogs? Is there a wild type for a dog that we could use as a standard? How would we

Mutations are
Consequences of Mutations
For an individual, the consequence of mutations may mean little or it may mean life or death. Some deleterious mutations are null or knock-out
Mutations and cancer
Mutations can affect either somatic cells or germ cells. Sometimes mutations occur in DNA repair genes, in effect compromising the cell's ability to fix other mutations that may arise. If,
Consequences of errors in replication, transcription and translation
Something key to think about:
Cells have evolved a variety of ways to make sure DNA errors are both detected and corrected,
Mutations as instruments of change
Mutations are how populations can adapt to changing environmental pressures
Mutations
Example: Antibiotic resistance
The bacterium E. coli is sensitive to an antibiotic called streptomycin, which inhibits protein synthesis by binding to the ribosome.
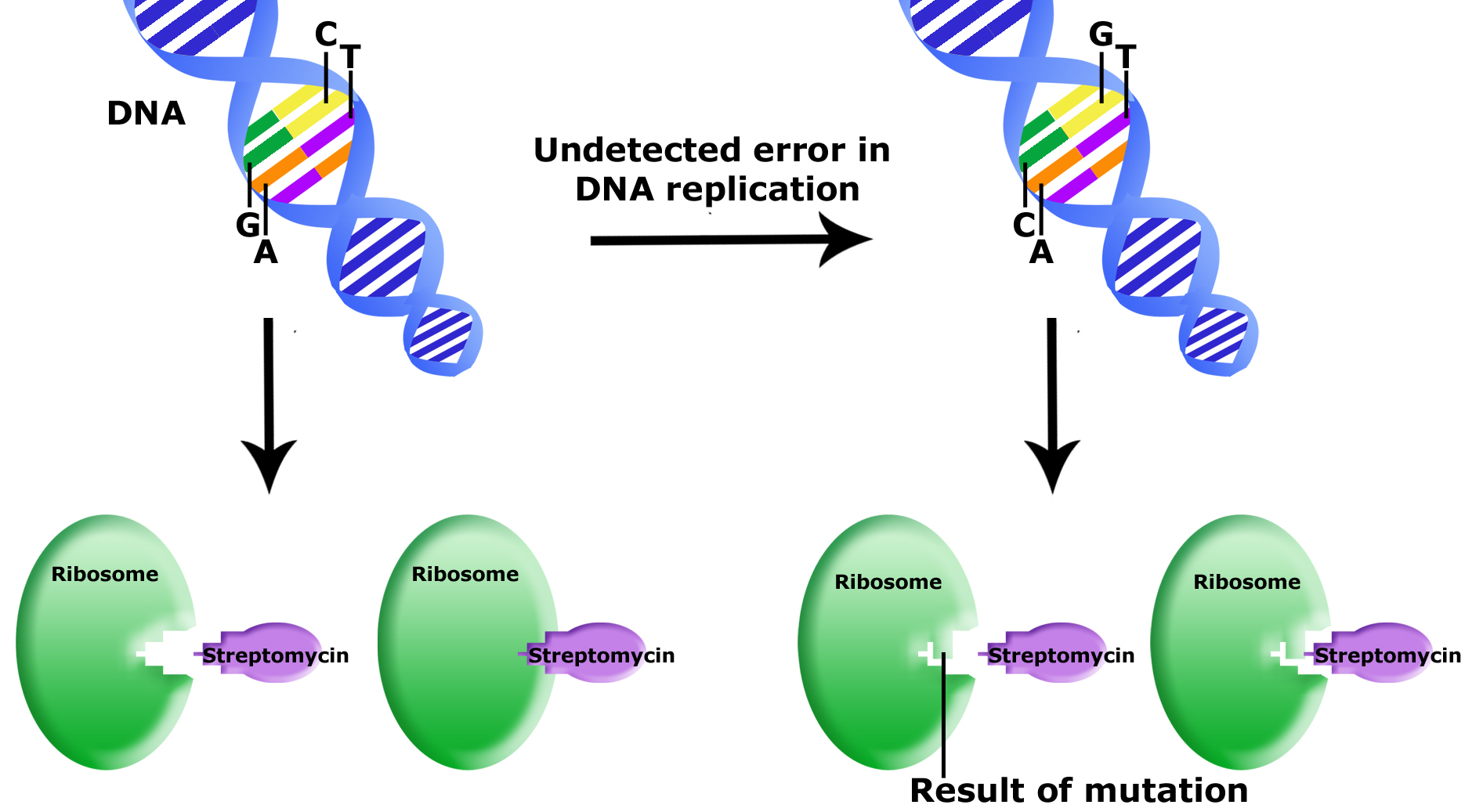
Uncorrected errors in DNA replication lead to mutation. In this example,
Source: Bis2A Team original image
An example: Lactate dehydrogenase
Lactate Dehydrogenase (LDH), the enzyme that catalyzes the reduction of pyruvate into lactic acid in fermentation, while virtually every organism has this activity, the corresponding enzyme and therefore gene differs immensely between humans and bacteria.
Possible NB Discussion Point
We can use comparative DNA sequence analysis to generate hypotheses about the evolutionary relationships between three or more organisms. One way to accomplish this is to compare the DNA or protein sequences of proteins found in each of the organisms we wish to compare. Let us, for example, imagine that we were to compare the sequences of lactate dehydrogenase (LDH) from three different organisms. The schematic below depicts the primary structures of LDH proteins from Organisms A, B, and C. The letters in the center of the proteins' line diagram represent amino acids at a unique position and the proposed differences in each sequences (Attribution:

Real-life Application:
As we have seen in the "Mutations and Mutants" module, changing even one nucleotide can have major effects on the translated product. Read more about an undergraduate's work on point mutations and GMOs here.
GLOSSARY
- induced mutation:
-
mutation that results from exposure to chemicals or environmental agents
- mutation:
-
variation in the nucleotide sequence of a genome
- mismatch repair:
-
type of repair mechanism in which
mismatched bases are removed after replication
- nucleotide excision repair:
-
type of DNA repair mechanism in which the wrong base, along with a few nucleotides upstream or downstream,
are removed
- proofreading:
-
function of DNA
pol in which it reads the newly added base before adding the next one
- point mutation:
-
mutation that affects a single base
- silent mutation:
-
mutation thatis not expressed
- spontaneous mutation:
-
mutation that takes place in the cellsas a result of chemical reactions taking place naturally without exposure to any external agent
- transition substitution:
-
when a purine
is replaced with a purine or a pyrimidineis replaced with another pyrimidine
- transversion substitution:
-
when a purine
is replaced by a pyrimidine or a pyrimidineis replaced by a purine
Eukaryotic Cell Cycle and Mitosis

The cell cycle is an orderly sequence of events used by biological systems to coordinate cell division. In eukaryotes, asexual cell division proceeds via a cell cycle that includes multiple spatially and temporally coordinated events. These include a long preparatory period called interphase, and a mitotic phase called M phase. Interphase is often further divided into distinguishable subphases called G1, S, and G2 phases. Mitosis is
Cell Cycle
In asexually reproducing eukaryotic cells, one “turn” of the cell cycle
Interphase
G1 Phase
The first stage of interphase
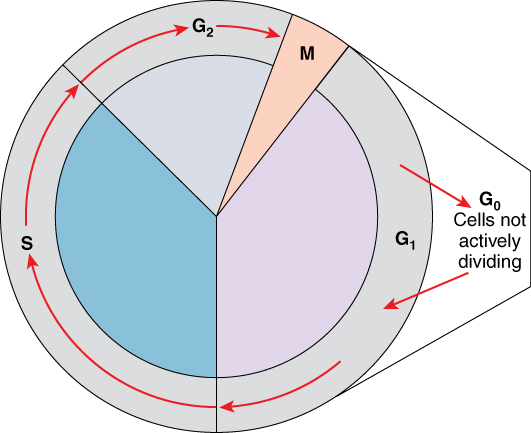
A cell moves through a series of phases in an orderly manner. During interphase, G1 involves cell growth and protein synthesis, the S phase involves DNA replication and the replication of the centrosome, and G2 involves further growth and protein synthesis. The mitotic phase follows interphase. Mitosis is nuclear division during which duplicated chromosomes
S Phase
Throughout interphase, nuclear DNA remains in a semi-condensed chromatin configuration. In S phase (synthesis phase), DNA replication results in the formation of two identical copies of each chromosome—sister chromatids—that
In cells using the organelles called centrosomes,
G2 Phase
During the G2 phase, or second gap, the cell replenishes its energy stores and synthesizes the proteins necessary for chromosome manipulation. Some cell organelles
G0 Phase
Not all cells adhere to the classic cell-cycle pattern in which a newly formed daughter cell immediately enters interphase, closely followed by the mitotic phase. Cells in the G0 phase are not actively preparing to divide. The cell is in a quiescent (inactive) stage, having exited the cell cycle. Some cells enter G0 temporarily until an external signal triggers the onset of G1. Other cells that never or rarely divide, such as mature cardiac muscle and nerve cells, remain in G0 permanently.
A Quick Aside: Structure of Chromosomes During the Cell Cycle
If we lay out the DNA from all 46 chromosomes end to end, it would measure approximately two meters; however, its diameter would be only 2
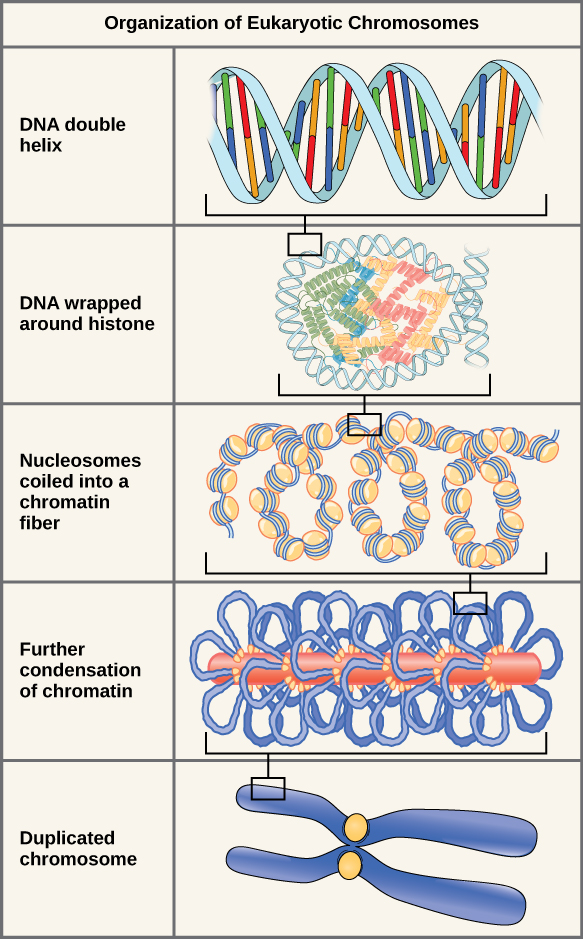
Double-stranded DNA wraps around histone proteins to form nucleosomes that appear like “beads on a string.”
Mitosis and Cytokinesis
During the mitotic phase, a cell undergoes two major processes. First, it completes mitosis, during which the contents of the nucleus
Note
The major phases of Mitosis are visually distinct from one another and

The stages of cell division oversee the separation of identical genetic material into two new nuclei, followed by the division of the cytoplasm.
Prophase
Prophase is the first phase of mitosis, during which the loosely packed chromatin coils and condenses into visible chromosomes. During prophase, each chromosome becomes visible with its identical partner (sister chromatid) attached, forming the familiar X-shape of sister chromatids. The nucleolus disappears early during this phase, and the nuclear envelope also disintegrates.
A major occurrence during prophase concerns a very important structure that contains the origin site for microtubule growth. Cellular structures called centrioles that serve as origin points from which microtubules extend. These tiny structures also play a very important role during mitosis. A centrosome is a pair of centrioles together. The cell contains two centrosomes side-by-side, which
Near the end of prophase there is an invasion of the nuclear area by microtubules from the mitotic spindle. The nuclear membrane has disintegrated, and the microtubules attach themselves to the centromeres that adjoin pairs of sister chromatids. The kinetochore is a protein structure on the centromere that is the point of attachment between the mitotic spindle and the sister chromatids.
Metaphase
Metaphase is the second stage of mitosis. During this stage, the sister chromatids, with their attached microtubules, line up along a linear plane in the middle of the cell. A metaphase plate forms between the centrosomes that
Anaphase
Anaphase is the third stage of mitosis. Anaphase takes place over a few minutes, when the pairs of sister chromatids
Telophase
Telophase is the final stage of mitosis.
Cytokinesis
Cytokinesis is the second part of the mitotic phase during which cell division
In cells such as animal cells that
In plant cells, a cleavage furrow is not possible because of the rigid cell walls surrounding the plasma membrane. A new cell wall must form between the daughter cells. During interphase, the Golgi apparatus accumulates enzymes, structural proteins, and glucose molecules prior to breaking up into vesicles and dispersing throughout the dividing cell. During telophase, these Golgi vesicles move on microtubules to collect at the metaphase plate. There, the vesicles fuse from the center toward the cell walls; this structure

In part (a), a cleavage furrow forms at the former metaphase plate in the animal cell.
Cell Cycle Check Points
It is essential that daughter cells be nearly exact duplicates of the parent cell. Mistakes in the duplication or distribution of the chromosomes lead to mutations that may pass forward to every new cell produced from the abnormal cell. To prevent a compromised cell from continuing to divide, there are internal control mechanisms that operate at three main cell cycle checkpoints at which
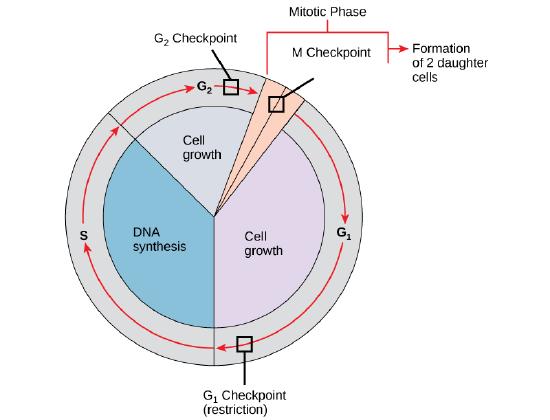
G1 Checkpoint
The G1 checkpoint determines whether all conditions are favorable for cell division to proceed into S phase where DNA replication occurs. The G1 checkpoint, also called the restriction point, is the point at which the cell irreversibly commits to the cell-division process. Besides adequate reserves and cell size, there is a check for damage to the genomic DNA at the G1 checkpoint.
G2 Checkpoint
The G2 checkpoint bars
M Checkpoint
The M checkpoint occurs near the end of the metaphase stage of mitosis.
Note
Watch what occurs at the G1, G2, and M checkpoints by visiting this animation of the cell cycle.
When the Cell Cycle gets out of Control
Most people understand that cancer or tumors
The process of a cell escaping its normal control system and becoming cancerous may
Homeostatic Imbalances: Cancer Arises from Homeostatic Imbalances
Cancer is
These two contrasting classes of genes, proto-oncogenes and tumor suppressor genes, are like the accelerator and brake pedal of the cell’s own “cruise control system,” respectively. Under normal conditions, these stop and go
A delicate homeostatic balance between the many proto-oncogenes and tumor suppressor genes delicately controls the cell cycle and ensures that only healthy cells replicate. Therefore, a disruption of this homeostatic balance can cause aberrant cell division and cancerous growths.