
Winter_2021_Bis2A_Facciotti_Reading_09_NB2
- Page ID
- 47831
Learning Objectives associated with Winter_2021_Bis2A_Facciotti_Reading_09
|
Introduction to bacterial and archaeal diversity
Perhaps bacteria may tentatively
Prokaryotes are single-celled organisms with neither a membrane-bound nucleus nor other lipid membrane-bound organelles.

Figure 1. Although bacteria and archaea are both described as prokaryotes,
Although bacteria and archaea share many morphological, structural, and metabolic attributes, there are many differences between the organisms in these two clades. The most notable differences are in the chemical structure and compositions of membrane lipids, the chemical composition of the cell wall, and the makeup of the information processing machinery (e.g., replication, DNA repair, and transcription).
Bacterial and archaeal diversity
Bacteria and archaea were on Earth long before multicellular life appeared. They are ubiquitous and have highly diverse metabolic activities. This diversity allows different species within clades to inhabit every imaginable surface where there is sufficient moisture. For example, some estimates suggest that in the typical human body, bacterial cells outnumber human body cells by about ten to one. Bacteria and archaea comprise most living things in all ecosystems. Certain bacterial and archaeal species can thrive in environments inhospitable for most other life. Bacteria and archaea, along with microbial eukaryotes, are also critical for recycling the nutrients essential for creating new biomolecules. They also drive the evolution of new ecosystems (natural or man-made).
The first inhabitants of Earth
So, when and where did life begin? What were the conditions on Earth when life began? What did LUCA (the Last Universal Common Ancestor), the predecessor to bacteria and archaea look like? While we don't know exactly when and how life arose and what it looked like when it did, we
The ancient atmosphere
Evidence
Note: The evolution of bacteria and archaea
How do scientists answer questions about the evolution of bacteria and archaea? Unlike
Scientists at the NASA Astrobiology Institute and at the European Molecular Biology Laboratory collaborated to analyze the molecular evolution of 32 specific proteins common to 72 species of bacteria. The model they derived from their data
The timelines of species divergence suggest that bacteria (members of the domain Bacteria) diverged from common ancestral species between 2.5 and 3.2 billion years ago, whereas archaea diverged earlier: between 3.1 and 4.1 billion years ago.
Microbial mats
Microbial mats (large
The first microbial mats likely harvested energy through redox reactions (discussed elsewhere) from chemicals found near hydrothermal vents. A hydrothermal vent is a breakage or fissure in the Earth’s surface that releases
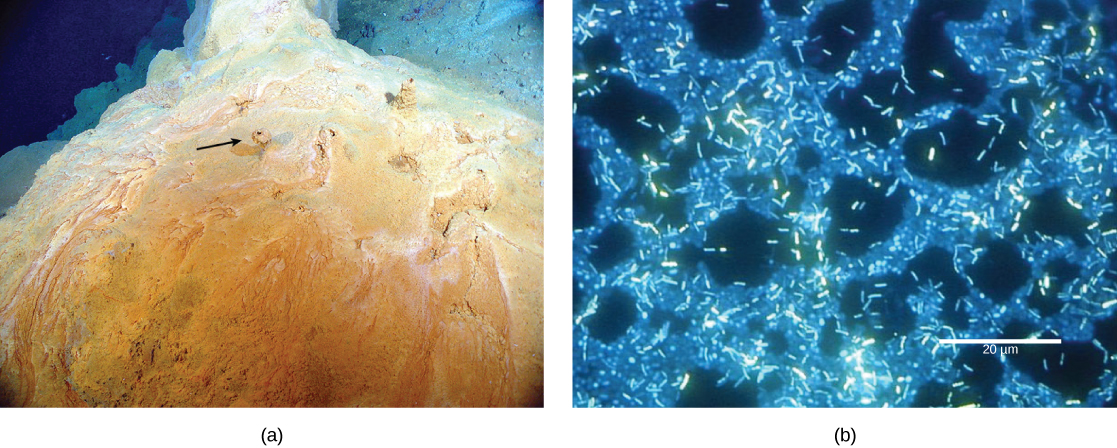
Figure 2. (a) This microbial mat, about one meter in diameter, grows over a hydrothermal vent in the Pacific Ocean in a region known as the “Pacific Ring of Fire.” Chimneys, such as the one shown by the arrow, allow gases to escape. (b) In this micrograph, bacteria within a mat
Stromatolites
A stromatolite is a sedimentary structure formed when minerals precipitate out of water

Figure 3. (a)
Bacteria and archaea are adaptable: life in moderate and extreme environments
Some organisms have developed strategies that allow them to survive harsh conditions. Bacteria and archaea thrive in a vast array of environments: some grow in conditions that would seem very normal to us, whereas others
Some bacteria and archaea
Possible NB Discussion  Point: How do extremophiles do it?
Point: How do extremophiles do it?
You just read that soil bacteria are able to survive through heat and droughts by forming dormant heat- and drought-resistant endospores. However, not all extremophiles form endospores as a means to survive their own harsh environmental conditions. Can you think of other strategies that other extremophiles might have developed? Choose a row in Table 1 (below) and try to brainstorm some creative survival mechanisms specific for that extremophile type!
| Extremophile Type | Conditions for Optimal Growth |
|---|---|
| Acidophiles | pH 3 or below |
| pH 9 or above | |
| Thermophiles | Temperature of 60–80 °C (140–176 °F) |
| Temperature of 80–122 °C (176–250 °F) | |
| Psychrophiles | Temperature of |
| Halophiles | Salt concentration of at least 0.2 M |
| High sugar concentration |
Figure
Footnotes
1. Battistuzzi, FU, Feijao, A, and Hedges, SB. A genomic timescale of prokaryote evolution: Insights into the origin of methanogenesis,
Cellular structure of bacteria and archaea
In this section, we will discuss the basic structural features of both bacteria and archaea. There are many structural, morphological, and physiological similarities between bacteria and archaea. As discussed in the previous section, these microbes inhabit many ecological niches and carry out a great diversity of biochemical and metabolic processes. Both bacteria and archaea lack a membrane-bound nucleus and membrane-bound organelles, which are hallmarks of eukaryotes.
While Bacteria and
Although bacteria and archaea come in a variety of shapes, the most common three shapes are

Figure 1. This figure shows the three most common shapes of bacteria and archaea: (a) cocci (spherical), (b) bacilli (rod-shaped), and (c)
Possible NB Discussion Point: Why are bacteria and archaea so tiny anyway?
Why are bacteria and archaea typically so small? What are the constraints that are keeping them microscopic in size (i.e., what is preventing from getting bigger?)? How then exactly does the relatively giant Thiomargarita namibiensis (which has a cell volume that is three million times the volume of an average bacteria and is visible to the naked eye) and other larger bacteria overcome these constraints? Think of
The bacterial and archaeal cell: common structures
Introduction to the basic cell structure
Bacteria and archaea are unicellular organisms, which lack internal membrane-bound structures that are disconnected from the plasma membrane, a phospholipid membrane that defines the boundary between the inside and outside of the cell. In bacteria and archaea, the cytoplasmic membrane also contains all membrane-bound reactions, including those related to the electron transport chain, ATP synthase, and photosynthesis. By definition, these cells lack a nucleus. Instead, their genetic material is located in a self-defined area of the cell called the nucleoid. The bacterial and archaeal chromosome is often a single covalently closed circular double-stranded DNA molecule. However, some bacteria have linear chromosomes, and some bacteria and archaea have more than one chromosome or small non-essential circular replicating elements of DNA called plasmids. Besides the nucleoid, the next common feature is the cytoplasm (or cytosol), the "aqueous," jelly-like region encompassing the internal portion of the cell. The cytoplasm is where the soluble (non-membrane-associated) reactions occur and contains the ribosomes, the protein-RNA complex where proteins are synthesized. Finally, many bacteria and archaea also have cell walls, the rigid structural feature surrounding the plasma membrane that helps provide protection and constrain the cell shape. You should learn to create a simple sketch of a general bacterial or archaeal cell from memory.

Figure 2. The features of a typical prokaryotic cell are shown.
Constraints on the bacterial and archaeal cell
One common, almost universal, feature of bacteria and archaea is that they are small, microscopic to be exact. Even the two examples given as exceptions, Epulopiscium fishelsoni and Thiomargarita namibiensis, still face the basic constraints all bacteria and archaea face; they simply found unique strategies around the problem. So what is the largest constraint when it comes to dealing with the size of bacteria and archaea? Think about what the cell must do to survive.
Some basic requirements
So what do cells have to do to survive? They need to transform energy into a usable form. This involves making ATP, maintaining an energized membrane, and maintaining productive NAD+/NADH2 ratios. Cells also need to be able to synthesize the appropriate macromolecules (proteins, lipids, polysaccharides, etc.) and other cellular structural components. To do this, they need to be able to either make the core, key precursors for more complex molecules or get them from the environment.
Diffusion and its importance to bacteria and archaea
Movement by diffusion is passive and proceeds down the concentration gradient. For compounds to move from the outside to the inside of the cell, the compound must be able to cross the phospholipid bilayer. If the concentration of a substance is lower inside the cell than outside and it has chemical properties that allow it to move across the cell membrane, that compound will energetically tend to move into the cell. While the "real" story is a bit more complex and will be discussed in more detail later, diffusion is one of the mechanisms bacteria and archaea use to aid in the transport of metabolites.
Diffusion can also be used to get rid of some waste materials. As waste products accumulate inside the cell, their concentration rises compared to that of the outside environment, and the waste product can leave the cell. Movement within the cell works the same way: compounds will move down their concentration gradient, away from where they are synthesized to places where their concentration is low and therefore may be needed. Diffusion is a random process—the ability of two different compounds or reactants for chemical reactions to interact becomes a meeting of chance. Therefore, in small, confined spaces, random interactions or collisions can occur more frequently than they can in large spaces.
The ability of a compound to diffuse depends on the viscosity of the solvent. For example, it is a lot easier for you to move around in air than in water (think about moving around underwater in a pool). Likewise, it is easier for you to swim in a pool of water than in a pool filled with peanut butter. If you put a drop of food coloring into a glass of water, it quickly diffuses until the entire glass has changed color. Now what do you think would happen if you put that same drop of food coloring into a glass of corn syrup (very viscous and sticky)? It will take a lot longer for the glass of corn syrup to change color.
The relevance of these examples is to note that the cytoplasm tends to be very viscous. It contains many proteins, metabolites, small molecules, etc. and has a viscosity more like corn syrup than water. So, diffusion in cells is slower and more limited than you might have originally expected. Therefore, if cells rely solely on diffusion to move compounds around, what do you think happens to the efficiency of these processes as cells increase in size and their internal volumes get bigger? Is there a potential problem to getting big that is related to the process of diffusion?
So how do cells get bigger?
As you've likely concluded from the discussion above, with cells that rely on diffusion to move things around the cell—like bacteria and archaea—size does matter. So how do you suppose Epulopiscium fishelsoni and Thiomargarita namibiensis got so big? Take a look at these links, and see what these bacteria look like morphologically and structurally: Epulopiscium fishelsoni and Thiomargarita namibiensis.
Based on what we have just discussed, in order for cells to get bigger, that is, for their volume to increase, intracellular transport must somehow become independent of diffusion. One of the great evolutionary leaps was the ability of cells (eukaryotic cells) to transport compounds and materials intracellularly, independent of diffusion. Compartmentalization also provided a way to localize processes to smaller organelles, which overcame another problem caused by the large size. Compartmentalization and the complex intracellular transport systems have allowed eukaryotic cells to become very large in comparison to the diffusion-limited bacterial and archaeal cells. We'll discuss specific solutions to these challenges in the following sections.
Membranes
Plasma membranes enclose and define the borders between the inside and the outside of cells. They are typically composed of dynamic bilayers of phospholipids into which various other lipid-soluble molecules and proteins have also been embedded. These bilayers are asymmetric—the outer leaf differing from the inner leaf in lipid composition and in the proteins and carbohydrates that are displayed to either the inside or outside of the cell. One major function of the outer cell membrane is to communicate the cell’s unique identity to other cells. The proteins, lipids, and sugars displayed on the cell membrane allow for cells to be detected by and to interact with specific partners.
Various factors influence fluidity, permeability, and various other physical properties of the membrane. These include temperature, the configuration of the fatty acid tails (some
Amoebae Hunting Video
Cellular membranes
A subgoal in our "build-a-cell" design challenge is to create a boundary that separates the "inside" of the cell from the environment "outside". This boundary needs to serve multiple functions that include:
- Act as a barrier by blocking some compounds from moving in and out of the cell.
- Be selectively permeable
in order to transport specific compounds into and out of the cell. - Receive, sense, and transmit signals from the environment to inside of the cell.
- Project "self" to others by communicating identity to other nearby cells.

Figure 1. The diameter of a typical balloon is 25cm and the thickness of the plastic of the balloon of around 0.25mm. This is a 1000X difference. A typical eukaryotic cell will have a cell diameter of about
Fluid mosaic model
The fluid mosaic model describes the dynamic movement of the
For some insight into the history of our understanding of the plasma membrane structure, click here.
It is sometimes useful to start our discussion by recalling the size of the cell membrane relative to the size of the entire cell

Figure 2. The fluid mosaic model of the plasma membrane describes the plasma membrane as a fluid combination of phospholipids, cholesterol, and proteins. Carbohydrates attached to lipids (glycolipids) and
The principal components of a plasma membrane are lipids (phospholipids and cholesterol), proteins, and carbohydrates. Carbohydrates are present only on the exterior surface of the plasma membrane and
Phospholipids
Phospholipids are major constituents of the cell membrane.
Make sure to note in Figure 3 that the phosphate group has an R group linked to one of the oxygen atoms. R is a variable commonly used in these types of diagrams to indicate that some other atom or molecule is bound at that position. That part of the molecule can be different in different phospholipids—and will impart some different chemistry to the whole molecule. At the moment, however, you are responsible for being able to recognize this type of molecule (no matter what the R group is) because of the common core elements—the glycerol backbone, the phosphate group, and the two hydrocarbon tails.

Figure 3. A phospholipid is a molecule with two fatty acids and a
Attribution:
When many phospholipids
Possible NB Discussion Point
Earlier in the course, we discussed the Second Law of Thermodynamics, which states that the overall entropy of the universe is always increasing. Apply this law in the context of the formation of the lipid bilayer membrane. How is it possible that the lipids are able to spontaneously arrange themselves into such an organized structure instead of scatter into a more disordered state? Or in other words -- if the second law holds true, then how exactly does the spontaneous lipid organization lead to increased entropy?

Figure 4. In the presence of water, some phospholipids will spontaneously arrange themselves into a micelle.
Source: Created by
Membrane proteins
Proteins make up the second major component of plasma membranes. Integral membrane proteins, as their name suggests, integrate completely into the membrane structure, and their hydrophobic membrane-spanning regions interact with the hydrophobic region of the phospholipid bilayer.
Some membrane proteins associate with only one half of the bilayer, while others stretch from one side of the membrane to the other, and are exposed to the environment on either side. Integral membrane proteins may have one or more transmembrane segments typically consisting of 20–25 amino acids. Within the transmembrane segments, hydrophobic amino acid variable groups arrange themselves to form a chemically complementary surface to the hydrophobic tails of the membrane lipids.
Peripheral proteins are found on only one side of the membrane, but never embed into the membrane. They can be on the intracellular side or the extracellular side, and weakly or temporarily associated with the membranes.

Figure 5. Integral membranes proteins may have one or more α-helices (pink cylinders) that span the membrane (examples 1 and 2), or they may have β-sheets (blue rectangles) that span the membrane (example 3). (credit: “Foobar”/Wikimedia Commons)
Carbohydrates
Carbohydrates are a third major component of plasma membranes. They are always found on the exterior surface of cells and are bound either to proteins (forming glycoproteins) or to lipids (forming glycolipids). These carbohydrate chains may consist of 2–60 monosaccharide units and can be either straight or branched. Along with peripheral proteins, carbohydrates form specialized sites on the cell surface that allow cells to recognize each other (one of the core functional requirements noted above.
Membrane fluidity
The integral proteins and lipids exist in the membrane as separate molecules and they "float" in the membrane, moving with respect to one another. The membrane is not like a balloon, however; because of the elastic properties of its plastic a balloon can easily grow and shrink its surface area without popping and while also maintaining the same rough circular shape. By contrast the plasma membrane is not able to withstand isotropic stretching or compression and can be easily popped when an imbalance of solute between inside and out causes water to rush in suddenly. A sudden loss of water will cause it to shrivel and wrinkle, dramatically changing the shape of the cell. it is fairly rigid and can burst if penetrated or if a cell takes in too much water and the membrane is stretched too far. However, because of its mosaic nature, a very fine needle can easily penetrate a plasma membrane without causing it to burst (the lipids flow around the needle point), and the membrane will self-seal when the needle is extracted.
Different organisms and cell types in multicellular organisms can tune fluidity of their membrane to be more compatible with specialized functions and/or in response to environmental factors. This tuning can be accomplished by adjusting the type and concentration of various components of the membrane, including the lipids, their degree of saturation, the lipids, their degree of saturation, the proteins, and other molecules like cholesterol. There are two other factors that help maintain this fluid characteristic. One factor is the nature of the phospholipids themselves. In their saturated form, the fatty acids in phospholipid tails are saturated with hydrogen atoms. There are no double bonds between adjacent carbon atoms, resulting in tails that are relatively straight. By contrast, unsaturated fatty acids do not have a full complement of hydrogen atoms on their fatty acid tails and therefore contain some double bonds between adjacent carbon atoms; a double bond results in a bend in the string of carbons of approximately 30 degrees.

Figure 6. Any given cell membrane will be composed of a combination of saturated and unsaturated phospholipids. The ratio of the two will influence the permeability and fluidity of the membrane. A membrane composed of completely saturated lipids will be dense and less fluid, and a membrane composed of completely unsaturated lipids will be very loose and very fluid.
Saturated fatty acids, with straight tails, are compressed by decreasing temperatures, and they will press in on each other, making a dense and fairly rigid membrane. Conversely, when unsaturated fatty acids are compressed, the “kinked” tails elbow adjacent phospholipid molecules away, maintaining some space between the phospholipid molecules. This “elbow room” helps to maintain fluidity in the membrane at temperatures at which membranes with high concentrations of saturated fatty acid tails would “freeze” or solidify. The relative fluidity of the membrane is particularly important in a cold environment. Many organisms (fish are one example) are capable of adapting to cold environments by changing the proportion of unsaturated fatty acids in their membranes in response to the lowering of the temperature.
Cholesterol
Animal cells have cholesterol, an additional membrane constituent that assists in maintaining fluidity. Cholesterol, which lies right in between the phospholipids in the membrane, tends to dampen the effects of temperature on the membrane.Cholesterol both stiffens and increases membrane fluidity, depending on the temperature. Low temperatures cause phospholipids to pack together more tightly, creating a stiffer membrane. In this case, the cholesterol molecules serve to space the phospholipids apart and prevent the membrane from becoming totally rigid. Conversely, higher temperatures contribute to phospholipids moving farther apart from each other and therefore a more fluid membrane, but cholesterol molecules in the membrane take up space and prevent the complete dissociation of phospholipids.
Thus, cholesterol extends, in both directions, the range of temperature in which the membrane is appropriately fluid and consequently functional. Cholesterol also serves other functions, such as organizing clusters of transmembrane proteins into lipid rafts.

Figure 7. Cholesterol fits between the phospholipid groups within the membrane.
Review of the components of the membrane
| The components and functions of the plasma membrane | |
|---|---|
| Component | Location |
| Phospholipid | Main fabric of the membrane |
| Cholesterol | Between phospholipids and between the two phospholipid layers of animal cells |
| Integral proteins (e.g., integrins) | Embedded within the phospholipid layer(s); may or may not penetrate through both layers |
| Peripheral proteins | On the inner or outer surface of the phospholipid bilayer; not embedded within the phospholipids |
| Carbohydrates (components of glycoproteins and glycolipids) | Generally attached to proteins on the outside membrane layer |
Archaeal membranes
One major difference setting archaea apart from eukaryotes and bacteria is their membrane lipid composition. Though eukaryotes, bacteria, and archaea all use glycerol backbones in their membrane lipids, Archaea use long isoprenoid chains (20-40 carbons in length, derived from the five-carbon lipid isoprene) that are attached via ether linkages to glycerol, whereas eukaryotes and bacteria have fatty acids bonded to glycerol via ester linkages.
The polar head groups differ based on the genus or species of the Archaea and consist of mixtures of glyco groups (mainly disaccharides), and/or phospho groups primarily of phosphoglycerol, phosphoserine, phosphoethanolamine or phosphoinositol. The inherent stability and unique features of archaeal lipids have made them a useful biomarker for archaea within environmental samples, though approaches based on genetic markers are now more commonly used.
A second difference between bacterial and archaeal membranes that is associated with some archaea is the presence of monolayer membranes, as depicted below. Notice that the isoprenoid chain is attached to the glycerol backbones at both ends, forming a single molecule consisting of two polar head groups attached via two isoprenoid chains.
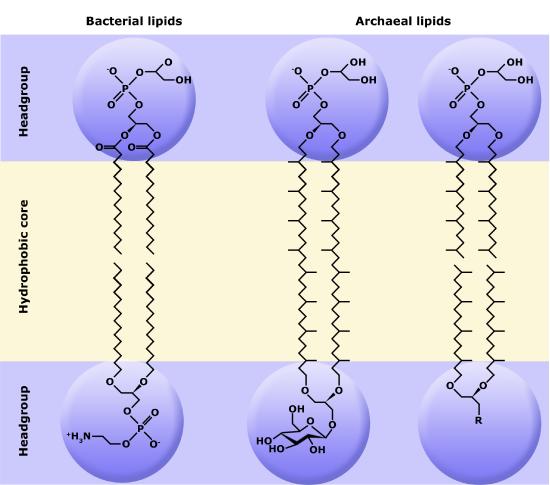
Figure 8. The exterior surface of the archaeal plasma membrane is not identical to the interior surface of the same membrane.
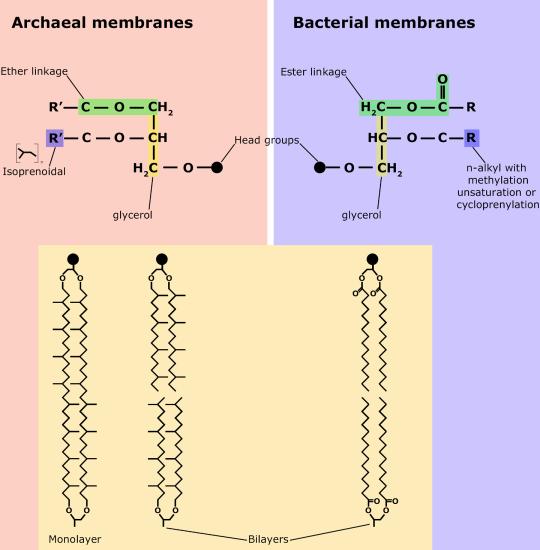
Figure 9. Comparisons of different types of archaeal lipids and bacterial/eukaryotic lipids
Transport across the membrane

Design challenge problem and subproblems
General Problem: The cell membrane must simultaneously act as a barrier between "IN" and "OUT" and control specifically which substances enter and leave the cell and how quickly and efficiently they do so.
Subproblems: The chemical properties of molecules that must enter and leave the cell are highly variable. Some subproblems associated with this are: (a) Large and small molecules or collections of molecules must be able to pass across the membrane. (b) Both hydrophobic and hydrophilic substances must have access to transport. (c) Substances must be able to cross the membrane with and against concentration gradients. (d) Some molecules look very similar (e.g.
Energy story perspective
We can consider transport across a membrane from an energy story perspective; it is a process
e.g. X
where in and out refer to inside the cell and outside the cell,
Selective permeability
One of the great wonders of the cell membrane is its ability to regulate the concentration of substances inside the cell. These substances include: ions such as Ca2+,
The membrane’s lipid bilayer structure provides the first level of control. The phospholipids pack tightly, and the membrane has a hydrophobic interior. This structure alone creates
Selective permeability of the cell membrane refers to its ability to differentiate between different molecules, only allowing some molecules through while blocking others. Some of this selective property stems from the intrinsic diffusion rates for different molecules across a membrane. A second factor affecting the relative rates of movement of various substances across a biological membrane is the activity of various protein-based membrane transporters, both passive and active, that will
Relative permeability
That different substances might cross a biological membrane at different rates should be relatively intuitive. There are differences in the mosaic composition of membranes in biology and differences in the sizes, flexibility, and chemical properties of molecules so
Membrane permeability coefficients
Below, we plot a variety of compounds
It is important that you can read and interpret the diagram below. The larger the coefficient, the more permeable the membrane is to the solute. For example,
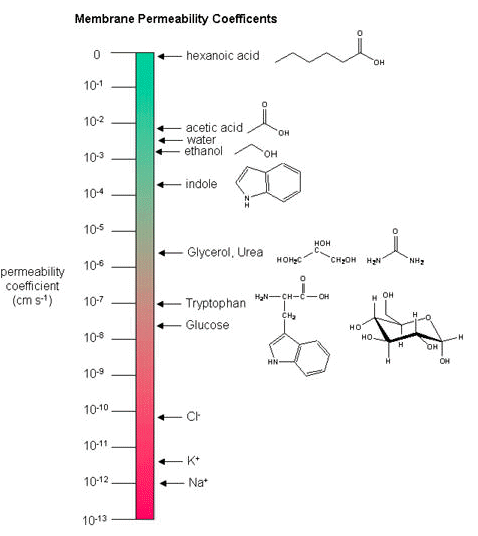
While there are certain trends or chemical properties that can
Osmosis
Osmosis is the movement of water through a semipermeable membrane according to the concentration gradient of water across the membrane, which is inversely proportional to the concentration of solutes. While diffusion transports material across membranes and within cells, osmosis transports only water across a membrane and the membrane limits the diffusion of solutes in the water. The
Mechanism
Osmosis is a special case of diffusion. Water, like other substances, moves from an area of high concentration to one of low concentration. An obvious question
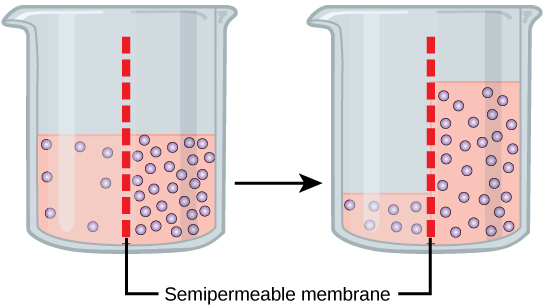
To illustrate this, imagine two full glasses of water. One has a single teaspoon of sugar in it, whereas the second one contains one-quarter cup of sugar. If the total volume of the solutions in both cups is the same which cup contains more water? Because the large amount of sugar in the second cup takes up much more space than the teaspoon of sugar in the first cup, the first cup has more water in it.
Returning to the beaker example, recall that it has a mixture of solutes on either side of the membrane. A principle of diffusion is that the molecules move around and will spread evenly throughout the medium if they can. However, only the material capable of getting through the membrane will diffuse through it. In this example, the solute cannot diffuse through the membrane, but the water can. Water has a concentration gradient in this system. Thus, water will diffuse down its concentration gradient, crossing the membrane to the side where it is less concentrated. This diffusion of water through the membrane—osmosis—will continue until the concentration gradient of water goes to zero or until the hydrostatic pressure of the water balances the osmotic pressure. Osmosis proceeds constantly in living systems.
Tonicity
Tonicity describes how an extracellular solution can change the volume of a cell by affecting osmosis. A solution's tonicity often directly correlates with the osmolarity of the solution. Osmolarity describes the total solute concentration of the solution. A solution with low osmolarity has a greater number of water molecules relative to the number of solute particles; a solution with high osmolarity has
Hypotonic solutions
Three terms—hypotonic, isotonic, and hypertonic—are used to relate the osmolarity of a cell to the osmolarity of the extracellular fluid that contains the cells. In a hypotonic situation, the extracellular fluid has
Hypertonic solutions
As for a hypertonic solution, the prefix hyper- refers to the extracellular fluid having a higher osmolarity than the cell’s cytoplasm; therefore, the fluid contains less water than the cell does. Because the cell has a relatively higher concentration of water, water will leave the cell.
Isotonic solutions
In an isotonic solution, the extracellular fluid has the same osmolarity as the cell. If the osmolarity of the cell matches that of the extracellular fluid, there will be no net movement of water into or out of the cell, although water will still move in and out. Blood cells and plant cells in hypertonic, isotonic, and hypotonic solutions take on characteristic appearances.
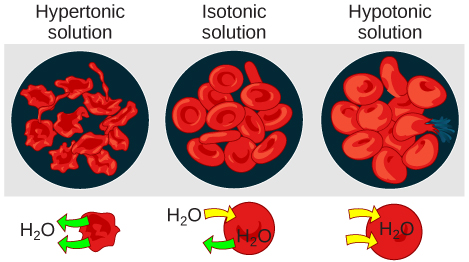
Figure 9. Osmotic pressure changes the shape of red blood cells in hypertonic, isotonic, and hypotonic solutions. (credit:
Possible NB Discussion Point
Of course there is such thing as drinking too little water... but is there such thing as drinking too much water? Discuss what you think happens when you drink an excessive amount of water -- what is happening at the level of the cell membrane? What is happening to the cell size? Is drinking too much water actually a health hazard? Predict what would happen if we changed the beverage to Gatorade instead of water.
Tonicity in living systems
In a hypotonic environment, water enters a cell, and the cell swells. In an isotonic condition, the relative concentrations of solute and solvent are equal on both sides of the membrane. There is no net water movement; therefore, there is no change in the
A red blood cell will burst, or lyse, when it swells beyond the plasma membrane’s capability to expand. Remember, the membrane resembles a mosaic, with discrete spaces between the molecules composing it. If the cell swells, and the spaces between the lipids and proteins become too large, and the cell will break apart.
In contrast, when excessive amounts of water leave a red blood cell, the cell shrinks. This has the effect of concentrating the solutes left in the cell, making the cytosol denser and interfering with diffusion within the cell. The cell’s ability to function will
Various living things have ways of controlling the effects of osmosis—a mechanism called osmoregulation. Some organisms, such as plants, fungi, bacteria, and some protists, have cell walls that surround the plasma membrane and prevent cell lysis in a hypotonic solution. The plasma membrane can only expand to the limit of the cell wall, so the cell will not lyse. In fact, the cytoplasm in plants is always slightly hypertonic to the cellular environment, and water will always enter a cell if water is available. This inflow of water produces turgor pressure, which stiffens the cell walls of the plant. In nonwoody plants, turgor pressure supports the plant. Conversely, if the plant
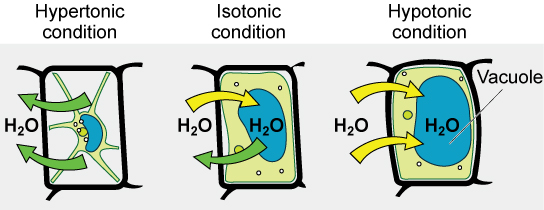

Tonicity is a concern for all living things. For example, paramecia and amoebas, which are protists that lack cell walls, have contractile vacuoles. This vesicle collects excess water from the cell and pumps it out, keeping the cell from bursting as it takes on water from its environment.
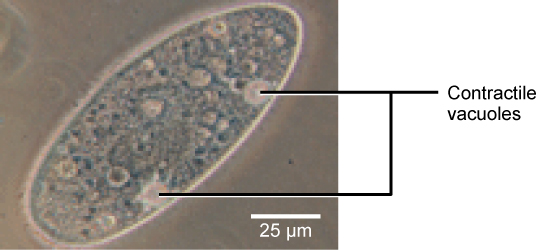
Figure 12. A paramecium’s contractile vacuole, here visualized using bright field light microscopy at 480x magnification, continuously pumps water out of the organism’s body to keep it from bursting in a hypotonic medium. (credit: modification of work by NIH; scale-bar data from Matt Russell)
Many marine invertebrates have internal salt levels matched to their environments, making them isotonic with the water in which they live. Fish, however, must spend approximately five percent of their metabolic energy maintaining osmotic homeostasis. Freshwater fish live in an environment that is hypotonic to their cells. These fish actively take in salt through their gills and excrete diluted urine to rid themselves of excess water. Saltwater fish live in the reverse environment, which is hypertonic to their cells, and they secrete salt through their gills and excrete highly concentrated urine.
In vertebrates, the kidneys regulate the amount of water in the body. Osmoreceptors are specialized cells in the brain that monitor the concentration of solutes in the blood. If the levels of solutes increase beyond a certain range, a hormone is released that retards water loss through the kidney and dilutes the blood to safer levels. Animals also have high concentrations of albumin, which is produced by the liver, in their blood. This protein is too large to pass easily through plasma membranes and is a major factor in controlling the osmotic pressures applied to tissues.