
2020_Spring_Bis2a_Facciotti_Lecture_25
- Page ID
- 27867
Learning Objectives Associated with 2020_Spring_Bis2a_Facciotti_Lecture_25
|
Examples of Bacterial Gene Regulation
This section describes two examples of transcriptional regulation in bacteria. These are presented as illustrative examples. Use these examples to learn some basic principles about mechanisms of transcriptional regulation. Be on the lookout in class, in discussion, and in the study-guides for extensions of these ideas and use these to explain the regulatory mechanisms used for regulating other genes.
Gene Regulation Examples in E. coli
The Role of the Promoter
The first level of control of gene expression is at the promoter itself. Some promoters recruit RNA polymerase and turn those DNA-protein binding events into transcripts more efficiently than other promoters. This intrinsic property of a promoter, it's ability to produce transcript at a particular rate,
UC Davis Undergraduate Connection:
A group of UC Davis students interested in synthetic biology used this idea to create synthetic
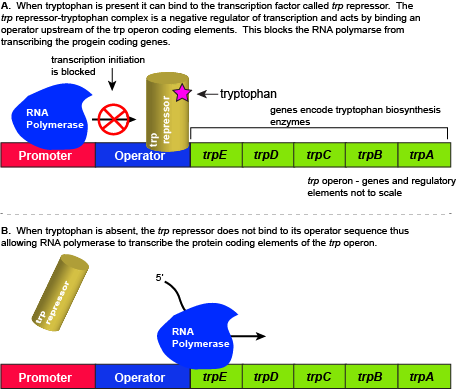
Example #1: Trp Operon
Logic for regulating tryptophan biosynthesis
E. coli, like all organisms, needs to either synthesize or consume amino acids to survive. The amino acid tryptophan is one such amino acid. E. coli can either import tryptophan from the environment (eating what it can scavenge from the world around it) or synthesize tryptophan de novo using enzymes encoded by five genes. These five genes
Organization of the trp operon
The five genes encoding tryptophan biosynthesis enzymes
A DNA sequence called an "operator"
A few more details regarding TF binding sites
We should note that the use of the term "operator" is limited to just a few regulatory systems and almost always refers to the binding site for a negatively acting transcription factor. Conceptually what you need to remember is that there are sites on the DNA that interact with regulatory proteins, allowing them to perform their appropriate function (e.g. repress or activate transcription). This theme will repeat universally across biology whether the "operator" term
While the specific examples you will be show depict TF binding sites in their known locations, these locations are not universal to all systems. Transcription factor binding sites can vary in location relative to the promoter. There are some patterns (e.g. positive regulators are often upstream of the promoter and negative regulators bind downstream), but these generalizations are not true for all cases. Again, the key thing to remember is that transcription factors (both positive and negatively acting) have binding sites with which they interact to help regulate the initiation of transcription by RNA polymerase.
The five genes that required to synthesize tryptophan in E. coli group next to each other in the
Attribution:
Regulation of the trp operon
When tryptophan is present in the cell: two tryptophan molecules bind to the
Since the transcription factor actively binds to the operator to keep the genes turned off, the
Possible NB Discussion  Point
Point
Suppose nature took a different approach to regulating the trp operon. Propose a method for regulating the expression of the
External link
Watch this video to learn more about the
Example #2: The lac operon
Rationale for studying the lac operon
In this example, we examine the regulation of genes encoding proteins whose physiological role is to import and assimilate the disaccharide lactose, the lac operon. The story of regulating lac operon is a common example used in many introductory biology classes to illustrate basic principles of inducible gene regulation. We describe this example second because it is, in our estimation, more complicated than the previous example involving the activity of a single negatively acting transcription factor.
As you go through this example, keep in mind the last point. For many Bis2a instructors, it is more important for you to learn the lac operon story and guiding principles than it is for you to memorize the logic table presented below. When this is the case, the instructor will usually let you know. These instructors often deliberately do NOT include exam questions about the lac operon. Rather, they will test you on whether you understood the fundamental principles underlying the regulatory mechanisms that you study using the lac operon example. If it's not clear what the instructor wants, ask.
The utilization of lactose
Lactose is a disaccharide composed of the hexoses glucose and galactose. We commonly encounter lactose in milk and some milk products. As one can imagine, the disaccharide can be an important food-stuff for microbes that can use its two hexoses. E. coli can use multiple different sugars as energy and carbon sources, including
However, there is a catch. Experiments conducted in the
The transcriptional regulators of the lac operon
The lac repressor - a direct sensor of lactose
As noted, the lac operon normally has very low to no transcriptional output in the absence of lactose. This is because of two factors: (1) the constitutive promoter strength for the operon is relatively low and (2) the constant presence of the LacI repressor protein negatively influences transcription. This protein binds to the operator site near the promoter and blocks RNA polymerase from transcribing the lac operon genes.
CAP protein - an indirect sensor of glucose
In E. coli, when glucose levels drop, the small molecule cyclic AMP (
In the lac operon, the
Note that the CAP-
Putting it all together: Inducing expression of the lac operon
For the lac operon to
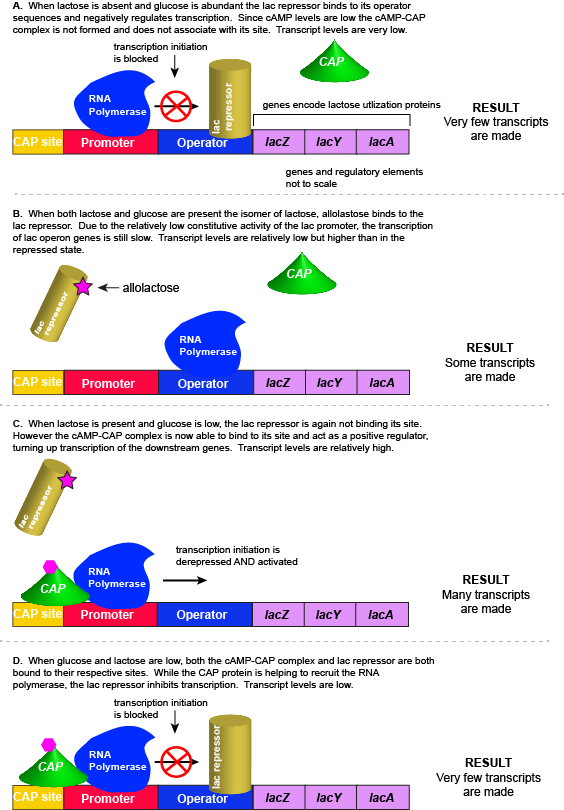
A more nuanced view of lac repressor function
The description of the lac repressor's function correctly describes the logic of the control mechanism used around the
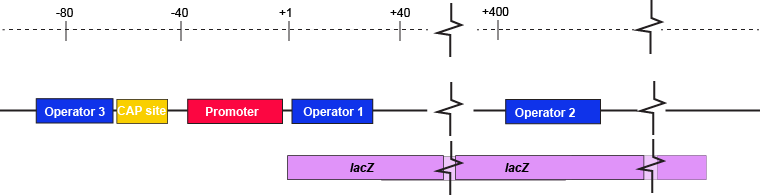
The lac operon regulatory region depicting the promoter, three lac operators, and CAP binding site.
Attribution:
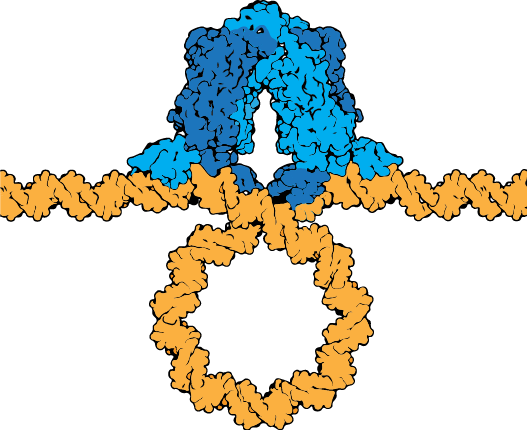
The lac repressor tetramer (blue) depicted binding two operators on a strand of looped DNA (orange).
Attribution:
Eukaryotic Gene Regulation
Regulation overview
As previously noted, regulation is all about decision making. Gene regulation, as a general topic, relates to
When we discussed the concept of strong and weak promoters, we introduced the idea that regulating the amount (number of molecules) of a transcript produced from a promoter in some unit of time might also be important for function. This should not be entirely surprising. For a protein-coding gene, the more transcript produced, the greater potential there is to make more protein. This might be important when making a lot of a particular enzyme is key for survival. In other cases, the cell needs only a little of a specific protein and making too much would be a waste of cellular resources.
In this section, we add to these themes by focusing on eukaryotic regulatory processes. Specifically, we examine - and sometimes re-examine - the multiple steps required to express genetic material in eukaryotic organisms in
Structural differences between bacterial and eukaryotic cells influencing gene regulation
The defining hallmark of the eukaryotic cell is the nucleus, a double membrane that encloses the cell's hereditary material. In order to efficiently fits the organism's DNA into the confined space of the nucleus, the DNA is first packaged and organized by protein into a structure called chromatin. This packaging of the nuclear material reduces access to specific parts of the chromatin. Some elements of the DNA are so tightly packed that the transcriptional machinery cannot access regulatory sites like promoters. This means that one of the first sites of transcriptional regulation in eukaryotes must be the control access to the DNA itself. Chromatin proteins can be subject to enzymatic modification that can influence whether they bind tightly (limited transcriptional access) or more loosely (greater transcriptional access) to a segment of DNA
The regulation of gene expression in eukaryotes also involves
The post-transcriptional enzymatic processing of RNA that occurs in the nucleus and the export of the mature
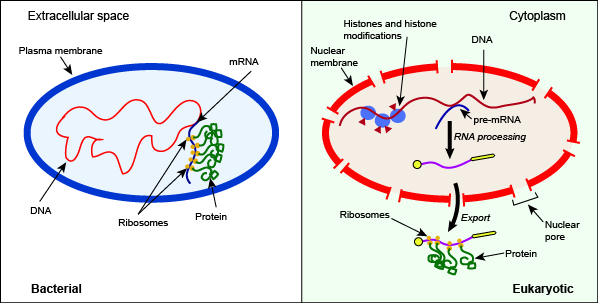
Depiction of some key differences between the processes of bacterial and eukaryotic gene expression. Note in this case
Attribution:
DNA Packing and Epigenetic Markers
The DNA in eukaryotic cells
DNA packing
The first level of organization, or packing, is the winding of DNA strands around histone proteins. Histones package and order DNA into structural units called nucleosomes, which can control the access of proteins to specific DNA regions. Under the electron microscope, this winding of DNA around histone proteins to form nucleosomes looks like small beads on a string. These beads (nucleosome complexes) can move along the string (DNA) to alter which areas of the DNA are accessible to transcriptional machinery. While nucleosomes can move to open the chromosome structure to expose a segment of DNA, they do so in a very controlled manner.
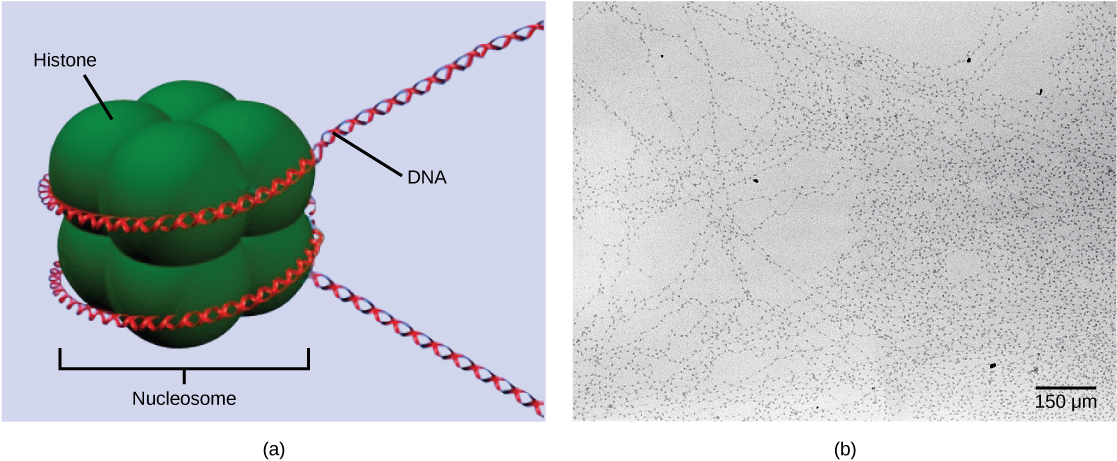
DNA folds around histone proteins to create (a) nucleosome complexes. These nucleosomes control the access of proteins to the underlying DNA. When viewed through an electron microscope (
Histone Modification
How the histone proteins move depends on chemical signals found on both the histone proteins and on the DNA. These chemical signals are chemical tags added to histone proteins and the DNA that tell the histones if a chromosomal region should be "open" or "closed". The figure below depicts modifications to histone proteins and DNA. These tags are not permanent, but may
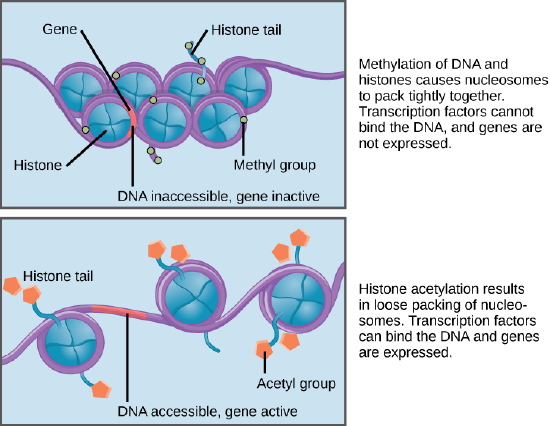
Nucleosomes can slide along DNA. When nucleosomes
Possible NB Discussion Point
In the later maturation phase of sperm cells, histones (containing high numbers of lysine amino acids) are replaced by protamines, which are small, nuclear proteins that are very rich in arginine amino acids. This process is said to be essential for sperm head condensation and DNA stabilization. Based on this information, what comparisons can you draw between protamines and histones? Why is it significant that there are high numbers of lysine and arginine in histones and protamines? For what reasons do you think protamines replace histones in sperm but not other cells?
DNA Modification
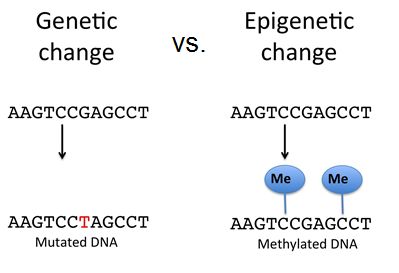
Epigenetic changes do not result in permanent changes in the DNA sequence. Epigenetic changes alter the chromatin structure (protein-DNA complex) to allow or deny access to transcribe genes. DNA modification such as methylation on cytosine nucleotides can either recruit repressor proteins that block RNA polymerase's access to transcribe a
Source:
Regulation of gene expression through chromatin remodeling
External link
View this video that describes how epigenetic regulation controls gene expression.
Eukaryotic gene structure and RNA processing
Eukaryotic gene structure
Many eukaryotic genes, particularly those encoding protein products,
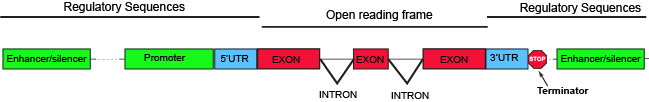
The parts of a typical discontinuous eukaryotic gene. Attribution:
Parts of a generic eukaryotic gene include familiar elements like a promoter and terminator. Between those two elements, the region encoding all the elements of the gene that have the potential to
The RNA species resulting from the transcription of these genes are also discontinuous and must therefore
5' capping
Like in bacterial systems, eukaryotic systems must assemble a pre-initiation complex at and around the promoter sequence to start transcription. The complexes that assemble in eukaryotes serve many of the same function as those in bacterial systems, but they are significantly more complex, involving many more regulatory proteins. This added complexity allows for greater regulation and for the assembly of proteins with functions that occur predominantly in eukaryotic systems. One of these additional functions is the "capping" of nascent transcripts.
In eukaryotic protein-coding genes, the RNA that is first produced
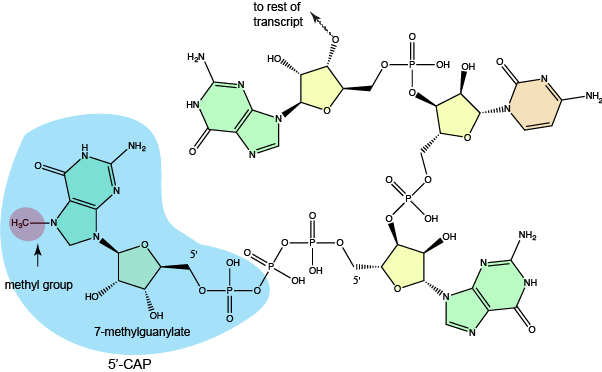
The structure of a typical 7-
Transcript splicing
Cells must process nascent transcripts into mature RNAs by joining exons and removing the intervening introns. They
One additional interesting outcome of alternative splicing is the introduction of stop codons that can, through a mechanism that seems to require translation, lead to the targeted decay of the transcript. This means that, besides the control of transcription initiation and 5'-capping, we can also consider alternative splicing as one of the regulatory mechanisms that may influence transcript abundance. The effects of alternative splicing are therefore potentially broad - from complete loss of function to novel and diversified function to regulatory effects.
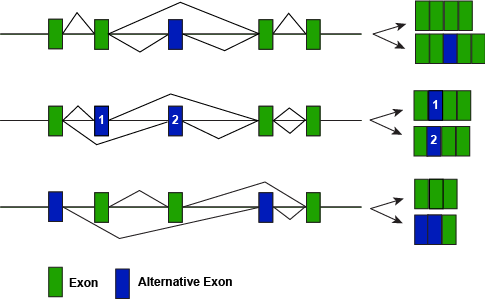
A figure depicting some different modes of alternative splicing illustrating how different splice variants can lead to different protein forms.
Attribution:
3' end cleavage and polyadenylation
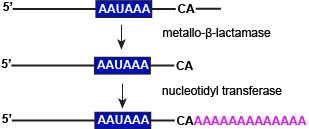
Attribution:
microRNAs
RNA Stability and microRNAs
Besides the modifications of the pre-RNA described above and the associated proteins that bind to the nascent and transcripts, there are other factors that can influence the stability of the RNA in the cell. One example are elements called microRNAs. The microRNAs, or miRNAs, are short RNA molecules that are only 21–24 nucleotides
Nuclear export
Nuclear export
Fully processed, mature
We know many additional details of the processes described above to some level of detail, but many more questions remain to
While we did not explicitly bring up the Design Challenge or Energy Story
Control of Protein Abundance
After a mRNA has been transported to the cytoplasm,
The initiation complex and translation rate
Like transcription,
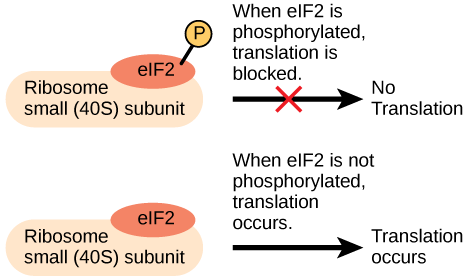
An increase in phosphorylation levels of
Chemical Modifications, Protein Activity, and Longevity
Not to
Chemical modifications can occur in response to external stimuli such as stress, the lack of nutrients, heat, or ultraviolet light exposure.
The addition of an ubiquitin group has another function - it marks that protein for degradation. Ubiquitin is a small molecule that acts like a flag
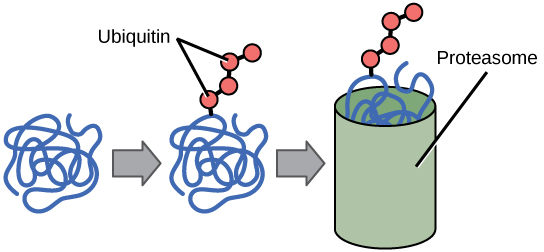
Proteins with ubiquitin tags are marked for degradation within the proteasome.
In conclusion, we see that gene regulation is complex and that