
2020_Spring_Bis2A_Facciotti_Lecture_13
- Page ID
- 27855
Learning Goals Associated with 2020_Spring_Bis2A_Facciotti_Lecture_13
|
Glycolysis: An overview

Organisms, whether unicellular or multicellular, need to find ways of getting at least two key things from their environment: (1) matter or raw materials for maintaining a cell and building new cells and (2) energy to help with the work of staying alive and reproducing. Energy and the raw materials may come from different places. For instance, organisms that primarily harvest energy from sunlight will get raw materials for building biomolecules from sources like CO2.
Glycolysis is the first metabolic pathway discussed in BIS2A;
The energy story and design challenge of glycolysis
Our investigation of glycolysis is a good opportunity to examine a biological process using both the energy story and the design challenge rubrics and perspectives.
The design challenge rubric will try to get you to think actively, and broadly and specifically, about why we are studying this pathway—what is so important about it? What "problems" does the evolution of a glycolytic pathway allow life to solve or overcome? We will also want to think about alternate ways to solve the same problems and why they may or may not have evolved. Later, we will examine a hypothesis for how this pathway—and other linked pathways—may have evolved, and thinking about alternative strategies for satisfying various constraints will come in handy then.
We ask you to think about glycolysis through the lens of an energy story in which you examine the 10-step process as a set of matter and energy inputs and outputs, a process with a beginning and an end. By taking this
So what is

| Enzyme | Step | ΔG/ |
ΔG°'/ |
|---|---|---|---|
| Hexokinase | 1 | -16.7 | |
| Phosphoglucose isomerase | 2 | -2.9 | 1.67 |
| Phosphofructokinase | 3 | -14.2 | |
| Fructose- |
4 | -0.23 | 23.9 |
| Triose phosphate isomerase | 5 | 2. |
7.56 |
| Glyceraldehyde 3-phosphate dehydrogenase | 6 | -1.29 | 6.30 |
| Phosphoglycerate kinase | 7 | 0.09 | -18.9 |
| Phosphoglycerate mutase | 8 | 0.83 | 4.4 |
| Enolase | 9 | 1.1 | 1.8 |
| Pyruvate kinase | 10 | -23.0 | -31.7 |
Overall, the glycolytic pathway comprises 10 enzyme-catalyzed steps. The primary input into this pathway is a single molecule of glucose, though we discover that other molecules may enter this pathway at various steps. We will focus our attention on (1) consequences of the overall process, (2) several key reactions that highlight important types of biochemistry and biochemical principles we will want to carry forward to other contexts, and (3) alternative fates of the intermediates and products of this pathway.
Note for reference that glycolysis is an anaerobic process. There is no requirement for molecular oxygen in glycolysis - oxygen gas is not a reactant in any of the chemical reactions in glycolysis. Glycolysis occurs in the cytosol or cytoplasm of cells. For a short (three-minute) overview YouTube video of glycolysis, click here.
First half of glycolysis: energy investment phase
We typically refer the first few steps of glycolysis as an "energy investment phase" of the pathway. This, however, doesn't make much intuitive sense (in the framework of a design challenge; it's not clear what
Step 1 of glycolysis:
The first step in glycolysis, shown below in Figure 2, is glucose being
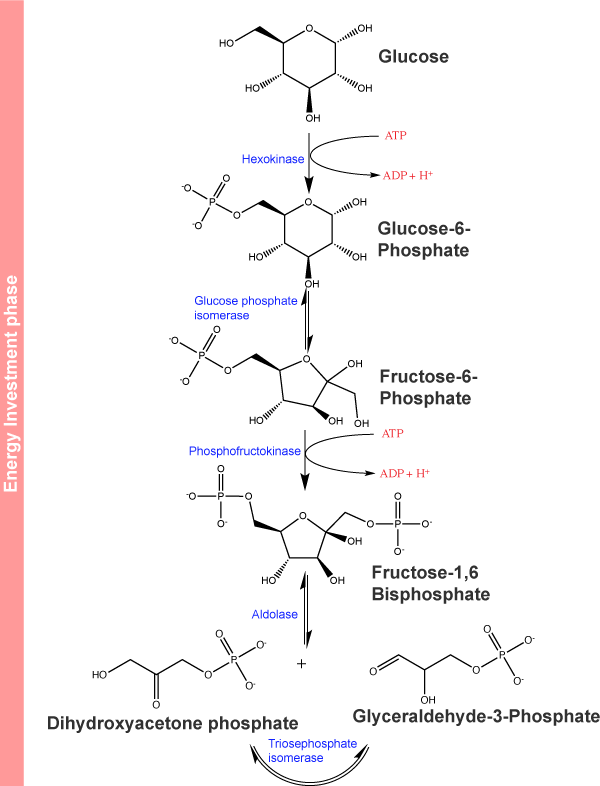
Figure 2. The first half of glycolysis
The paragraph above states that the enzyme hexokinase has "broad specificity." This means that it can
The conversion of glucose to the negatively charged glucose 6-phosphate significantly reduces the likelihood that the phosphorylated glucose leaves the cell by diffusion across the hydrophobic interior of the plasma membrane. It also "marks" the glucose in a way that tags it for several

Figure 3. Note that this figure shows that glucose 6-phosphate can, depending on cellular conditions,
As Figure 3 shows, glycolysis is but one fate for glucose 6-phosphate (G6P). Depending on cellular conditions, G6P may
Step 2 of glycolysis:
In the second step of glycolysis, an isomerase
Step 3 of glycolysis:
The third step of glycolysis is the phosphorylation of fructose 6-phosphate,
Step 4 of glycolysis:
In the fourth step in glycolysis, an enzyme, fructose-
Second half: energy payoff phase
If viewed in the absence of other metabolic pathways, glycolysis has so far cost the cell two ATP molecules and produced two small, three-carbon sugar molecules: dihydroxyacetone phosphate (DAP) and glyceraldehyde 3-phosphate (G3P). When viewed in a broader context, this investment of energy to produce a variety of molecules that can
Both DAP and G3P can proceed through the second half of glycolysis. We now examine these reactions.

Figure 4. The second half of glycolysis
Step 5 of glycolysis:
In the fifth step of glycolysis, an isomerase transforms the dihydroxyacetone phosphate into its isomer, glyceraldehyde 3-phosphate. The six-carbon glucose has therefore now
Step 6 of glycolysis:
The sixth step is key and one from which we can now leverage our understanding of the several chemical reactions that we've studied so far. If
It is important to note that this reaction depends upon the availability of the oxidized form of the electron carrier, NAD+. If we consider that there is a limiting pool of NAD+, we can then conclude that the reduced form of the carrier (NADH) must continuously oxidize back into NAD+ to keep this step going. If NAD+ is not available, the second half of glycolysis slows down or stops.
Possible NB Discussion  Point
Point
Can you write an energy story for Step 6 of glycolysis (the reaction
Step 7 of glycolysis:
In the seventh step of glycolysis, catalyzed by phosphoglycerate kinase (an enzyme named for the reverse reaction), 1,3-
Step 8 of glycolysis:
In the eighth step, the remaining phosphate group in 3-phosphoglycerate moves from the third carbon to the second carbon, producing 2-phosphoglycerate (an isomer of 3-phosphoglycerate). The enzyme
Step 9 of glycolysis:
Enolase
Step 10 of glycolysis:
The last step in glycolysis
Outcomes of glycolysis
Here are a few things to consider:
One of the clear outcomes of glycolysis is the biosynthesis of compounds that can enter
If glycolysis runs long enough, the constant oxidation of glucose with NAD+ can leave the cell with a problem: how to regenerate NAD+ from the two molecules of NADH produced. If the cell does not regenerate NAD+, nearly all the cell's NAD+ will transform into NADH. So how do cells regenerate NAD+?
Pyruvate
Possible NB Discussion Point
To some, that glycolysis is such a complex, multi-step pathway may seem counter-intuitive: “Why wouldn’t evolution lead to a *simpler* way to extract energy from food since energy is an important requirement for life?” Explain the necessity/advantage of having glucose get broken down in many steps.
Substrate-level phosphorylation (SLP)
The simplest route to synthesize ATP is substrate-level phosphorylation. ATP molecules

In this reaction, the reactants are a phosphorylated carbon compound called G3P (from step 6 of glycolysis) and an ADP molecule, and the products are 1,3-BPG and ATP. The transfer of the phosphate from G3P to ADP to form ATP in the active site of the enzyme is substrate-level phosphorylation. This occurs twice in glycolysis and once in the TCA cycle (for a subsequent reading).