
2020_SS1_Bis2A_Facciotti_Reading_16
- Page ID
- 34020
Learning objectives associated with 2020_SS1_Bis2A_Facciotti_Reading_16• Describe the mechanism of allosteric regulation and provide examples of how this mechanism can be used to tune the flow of molecules in a metabolic pathway. |
An introduction to metabolic regulation
Metabolic pathways
We can describe cellular metabolic pathways as sets of interconnected enzyme-catalyzed chemical reactions. Together these pathways form complex networks that collectively guide the flow of molecules and energy throughout a cell by breaking down, building, and rearranging the atoms and molecules of nature into their many different forms.
The collective activity of metabolic pathways allows organisms to eat food (e.g. atoms and molecules) from their environments and to convert that food at a molecular level into the specific forms of matter the organism needs to support life. Metabolic pathways also allow organisms to harvest energy from the environment and to move that energy to places in their cells that need energy to get work done.
Understanding how metabolic pathways help biological systems reorganize matter and energy is therefore fundamental to our understanding of biology from the smallest cellular scale to the functioning of connected global ecosystems and their interactions with the non-living environment.
Metabolic decision making, sensors and switches
Many living systems have complex metabolic pathways that enable them to make and process many different types of molecules. While these complex pathways likely provide an organism with selective advantage not all pathways need to be functioning at the same levels all of the time.
If, for instance, a unicellular organism is able to find and eat plenty of the amino acid tryptophan from its environment, the organism’s metabolic pathways that synthesize tryptophan do not need to be highly active. Running metabolic pathways that don’t need to be active wastes both the atoms that pass through the pathways and the energy used in the creation of molecules that aren’t needed at a particular time. Moreover, since most living systems do not exist in resource-rich environments, atoms and energy must be carefully distributed to cellular functions that need them most, only when they are needed. Resources can then be converted into other forms when they are needed elsewhere in the cell.
To be competitive, the cell must be frugal with resources and thus control how and when different metabolic pathways are active. Controlling the flow of materials through metabolic pathways requires mechanisms for decision making - we use the anthropomorphic word “decision” to imply that the cell seemingly knows how to make choices, not to suggest that it has a brain. For example, the cell in the example above needs a way to know that tryptophan is in abundance and then to use that information to switch off the biosynthetic pathway for tryptophan biosynthesis. Concomitantly, the cell can choose to use the metabolic precursors for tryptophan biosynthesis for another purpose, activating enzymes in other pathways that can use those same precursor molecules. This kind of sensing and control is required to simultaneously coordinate the flow of biomolecules and energy throughout all of cellular metabolism. This means that there must be molecular sensors and switches for controlling metabolism distributed throughout the cell.

Figure 1. Overview of the major connections between core metabolic pathways in eukaryotes.
Attribution:Chakazul / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0) <https://upload.wikimedia.org/wikipedia/commons/6/6e/Metabolic_Metro_Map.svg>
Our daily lives are full of similar experiences and concepts that can inform how we think about metabolic control in biology. Our homes have all sorts of sensors and switches for managing energy. We have light switches in every room to control the flow of electricity and conversion of energy through light bulbs. Our stoves have switches on them to control energy use on each burner. Thermostats in the home sense temperature and use that information to adjust the activity of our air conditioners and/or heaters. Some of these switches are bistable, they turn things fully “on” or “off” (e.g. most light switches). Other switches are continuous (e.g. dial switches on a stove, the volume knob on the radio). They allow the user to control energy use along a gradient.
The hierarchical nature of regulatory systems
Another useful concept to draw from looking at how we manage energy in our homes is to note the hierarchical nature of the control systems we have engineered for our living spaces. Nearly all homes/apartments have circuit breakers that can help control the flow of power from an energy provider into the home. In a circuit breaker box one usually finds a master switch that can turn power on or off for the entire home. One can usually also find switches that control power to individual rooms or parts of the house (e.g. the kitchen, the bathrooms). Within each room, light switches mounted on the wall typically allow the resident to control energy flow to all or just some of the outlets in the wall. Finally, each device plugged into the wall has its own “on”/“off” switch. Many devices, like a radio, have additional switches on them that allow individualized energy use management. As we develop our understanding of regulation in biology, we will see that Nature has also evolved conceptually similar hierarchical control structures (though typically more interconnected) to help manage metabolism and various other cellular functions. Some signaling molecules target and regulate very specific control points - a single enzyme. Other molecular signals target many control points simultaneously (many enzymes can be coordinately regulated). The interplay between specific control signals and broader control signals creates the ability to respond to big changes quickly and also to fine tune responses in the cell in a very detailed way.
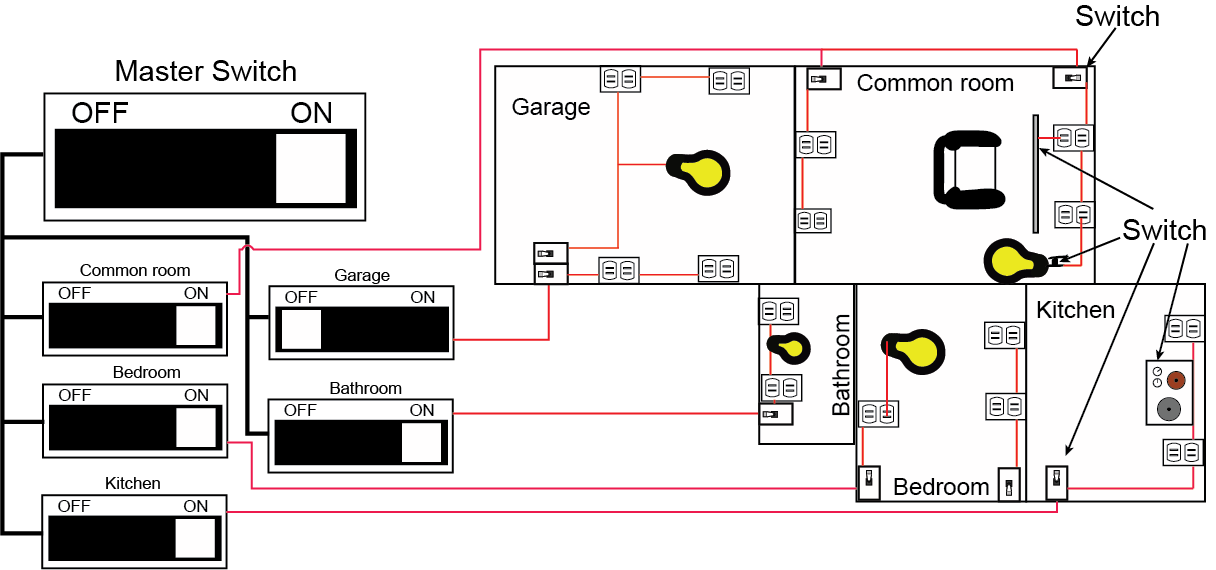
Figure 2. A schematic of a household wiring scheme showing hierarchical control of power to rooms, lights and devices. The master switch can turn off all power. Sub-switches can control power to individual rooms. Meanwhile, room-level switches can control power to parts of the room. Finally, switches on devices can control power at an individual device level. This serves as an analogy for some features of metabolic and cellular regulation.
Attribution: Original work - Marc T. Facciotti
The Who, What, When, Where, Why and How of Regulation
We structure the rest of this discussion of metabolic regulation by using a formalism often associated with journalism. We examine answers to the questions who, what, when, where, why and how <https://en.wikipedia.org/wiki/Five_Ws>.
Why regulate?
To some degree, we have made the argument about why regulation is necessary above. Largely, regulation in metabolism is involved in making decisions about how to best manage limited resources - the atoms that make up biomolecules, the biomolecules themselves, and the energy needed to build new molecules and/or carry out cellular functions. Good decision-making leads to more efficient resource utilization and by consequence we suspect higher evolutionary fitness.
Who to regulate?
When we consider the regulation of metabolism and metabolic pathways, the key control points are the enzymes that catalyze individual reactions. In the simplest terms, increasing the abundance of a specific enzyme and/or increasing its activity (how fast it catalyzes a reaction) can increase the flow of molecules through that step of a pathway. By contrast, decreasing the abundance of an enzyme and/or lowering its activity will reduce the amount of material flowing through that step in a pathway.
Where should regulation be exercised?
If enzymes and their relative abundance are good targets of regulation we can now ask whether some steps in metabolic pathways are more important to regulate than others. The answer to this question is yes and can be largely summarized by finding enzymes that belong to one or more of three classes: (a) enzymes that catalyze steps at branch points in a pathway; (b) enzymes that catalyze rate-limiting reactions; and (c) enzymes that catalyze so-called irreversible reactions (i.e. reaction with a large negative ∆G). We’ll examine the logic behind each.
Enzymes that catalyze reactions at branch points: Branch points in metabolic reactions are points in a metabolic pathway in which a compound can be used as a substrate in two or more different biochemical reactions. The cell must “decide” which of the two or more pathways to direct the biomolecule to. This is no different than the everyday experience we each time we encounter an intersection in the road we’re traveling on - we must decide whether to continue straight or turn down a different path. We can’t do both at the same time. Another everyday example might be in the construction of a water distribution pipeline in a garden with two or more planting beds. Depending on what is planted in each bed the gardener may wish to tune the flow of water to each bed differently and would do so by opening and closing valves at the branch points of the watering system.
Enzymes that catalyze rate-limiting steps in a pathway: A rate-limiting step in a metabolic pathway is defined as the reaction that determines how fast the overall pathway overall can convert input into output. This is almost always the slowest step in the pathway. Regulating this step by either slowing or speeding flow of metabolites through it can change the flow for the entire pathway. By analogy, you can think of a 5-lane highway that needs to constrict to 3 lanes over a bridge. While the highway may widen back up to 5 lanes after the bridge, the maximum rate of traffic flow before the bridge and after the bridge is limited by how many cars can get across the bridge at any one time. Narrowing or widening the bridge will have a major impact on traffic flow both before and after.
Enzymes that catalyze irreversible reactions: Reactions that have a large negative ∆G are often called irreversible reactions. These reactions are also often considered to be “commitment” steps in a biochemical pathway because it is difficult to reverse the reaction once it is done. Typically, different enzymes and external energy sources are required to run the reaction in the endergonic direction. We can also draw upon an analogy here to understand why these are important decision points by thinking about decisions in life that are hard to take back once they are made. Consider, for instance, the act of buying a used car or other equivalent item of value that you might need, or care to have. Few people would consider buying such an item sight unseen and without asking questions, doing some research, and maybe even getting a professional opinion on the condition of the car or other item. If the car/item is still functional and well-maintained the purchase could work out fantastically well. On the other hand, if the car/item has some defects or in need of repair, the purchase may not end well at all. The decision to buy the car/item must be carefully considered BEFORE the purchase. After you hand over the money there is typically no opportunity to changing your mind – the sale if final - and you will need to live with the consequences of that decision, good or bad. The same idea applies to reactions with large negative ∆G. Once the decision has been made to catalyze the reaction, that decision cannot be easily reversed. Therefore, the cell must evolve mechanisms that help make these decisions carefully on the basis of good information.
In many metabolic pathways, particularly those that are as interconnected to other pathways as glycolysis, the picture is - of course - more complicated as each these three types of control points are each used for regulation and it is too simplistic to simply state that there is a single point in the pathway that controls flow. Rather, it is the combination and interplay of regulatory sites and regulatory mechanisms that ultimately determines flow into, through, and out of the pathway.
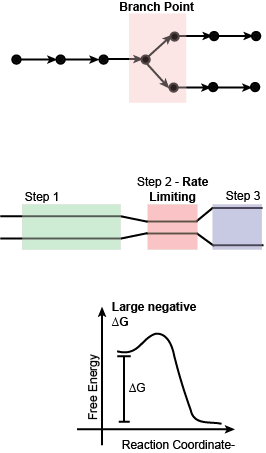
Figure 3. Schematic of three important types of steps in metabolic pathways that are key targets of regulation.
Attribution: Original work - Marc T. Facciotti
When to regulate?
A cell is always regulating the flow of molecules through its metabolic pathways. Understanding when to change the regulation of different metabolic pathways is the critical issue. When does the organism start to run glycolysis? When does it ramp up nucleotide biosynthesis and turn down amino acid biosynthesis? The short answer is that pathways are up-regulated when they are needed and down-regulated when they aren’t. But how does the organism know when something is needed and something else isn’t? The answer to this question is a bit more involved than what we can discuss in the time and space provided here. However, we can intuit that whatever these processes are, they must involve the sensing of environmental and/or cellular information. Something must be measured.
What to measure?
In the context of metabolic pathways, what information could be measured to help a cell make decisions about whether to up- or down-regulate flow through a metabolic pathway? Since metabolic pathways are in “the business” of consuming input molecules and converting them into some other product, it might be reasonable to expect that knowing whether there is enough product in the cell already (in which case the pathway doesn’t need to run) or whether there is enough of the original substrate around to feed the pathway could be useful information to know for regulation. Other cellular information of relevance to metabolism that might be useful to know about when deciding on whether to up- or down-regulate a pathway is the general level of usable energy (e.g. the balance between levels of ATP, ADP, and AMP) and the availability of reduced and oxidized electron carriers (e.g. the balance between NADH and NAD+). Clearly, in some cases the levels and balance between other molecules will also be important.
How can the cell use molecular information to make metabolic decisions?
Above we suggested some rationale about the regulation of metabolic pathways. We proposed that metabolic pathways must be regulated to manage cellular resources (why) and that this regulation should happen in response to changing cellular needs (when). We propose that regulation happens when branch points, irreversible reactions, and rate-limiting steps are found (where). Furthermore, we suggest that regulation happen by controlling the abundance and/or activity of the enzymes that catalyze reactions in a pathway (who). Finally, we posit that knowing the abundance of pathway inputs and products as well as general indicators of cellular energy and redox stores (what) is generally useful for cellular decision-making.
This leaves us to answer the final question. How? How can a cell use abundance of cellular molecules to inform enzymes catalyzing key steps in metabolic pathways about whether they are needed more or less when cellular conditions change? The complete answer to this question is, like seemingly everything else in biology, multifaceted, involving different targets of regulation (e.g. the genes encoding an enzyme to the enzyme itself) and different mechanisms. Here we briefly discuss one mechanism that links back to an earlier lesson on proteins; allosteric regulation.
Allosteric regulation involves the binding of molecules to allosteric sites on an enzyme that are by definition not in the active site. This binding alters the protein structure and can cause in different instances both either up or down regulation of enzyme activity. Often, enzymes that are the subject of allosteric regulation can be influenced by more than one ligand. By binding different phosphorylation states of ATP (i.e. AMP, ADP, and ATP) at allosteric sites enzymes can measure the ratios of ATP to its other forms and thus assess the energy “status” of the cell.
Examples of metabolic regulation by allosteric binding
Reactions with large negative ∆G
An early intermediate step of glycolysis, the phosphorylation of fructose-6-phosphate by ATP to yield fructose-1,6-bisphosphate and ADP, has an approximate ∆G of -19kJ/mol under cellular conditions. This is one of three largest, by comparison to all other reactions in the pathway, drops in free energy in the glycolytic pathway. In mammalian systems, the enzyme catalyzing this reaction, phosphofructokinase, is both positively and negatively regulated by multiple small molecules. ATP, citrate, and phosphoenolpyruvate (PEP) have been found to bind allosteric sites on the enzyme and lead to a lowering of enzyme activity. AMP, ADP, and fructose-2,6-bisphosphate (a product created by another enzyme) each increase phosphofructokinase activity. A number of other small molecules have also been found to influence enzyme activity in the test tube, but these are typically considered to play an insignificant role in the living system.
Given the central role of the glycolytic pathway in both energy harvesting and the creation of key precursors for other metabolic pathways, it is perhaps not surprising to have this enzyme’s activity influenced by a measure of the energy status of the cell (the ATP/AMP ratio) and by molecules in connected pathways (PEP and citrate). It is perhaps easiest to understand that when the enzyme senses low energy levels in the cell, when the ATP/AMP ratio is low, that the enzyme committing sugar to enter the energy extraction phase of glycolysis should become more active and that the converse should be true when ATP is abundant.

Figure 4. The enzyme phosphofructokinase is regulated allosterically both positively and negatively by numerous small molecules indicators of cellular energy state and of key concentrations of pathway intermediates.
Attribution: Original work - Marc T. Facciotti
Branch Points
An example of branch point regulation can be found in the pathways leading to the biosynthesis of aromatic amino acids (i.e. Phenylalanine, Tyrosine, and Tryptophan). The synthesis of each of these three amino acids begins with the production of the compound Chorismate. This compound can then be taken down two independent pathways. The first leads to Phenylalanine and Tyrosine biosynthesis while the second leads to Tryptophan biosynthesis. The enzyme catalyzing the first step in the path towards Phenylalanine and Tyrosine, Chrosimate mutase, is negatively regulated by the two products of the pathway and activated by Tryptophan, the product of the second pathway. Meanwhile, the enzyme Anthranilate Synthase is negatively regulated by the final product of its pathway, Tryptophan. The feedback of end-product levels on the enzymes responsible for catalyzing the synthesis of these three amino acids allow the cell to decide how to best utilize the stock of Chorismate and make that decision at the one of the core branch points for its use.
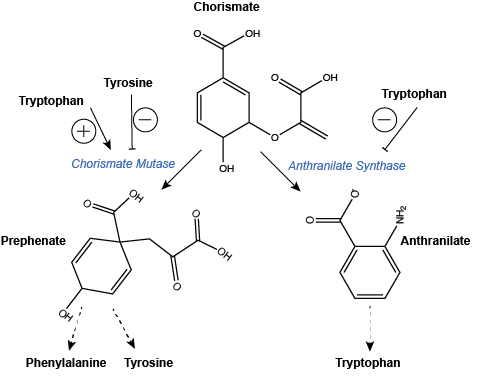
Figure 5: Regulation of branchpoint enzymes in the biosynthesis of aromatic amino acids. Dashed arrows indicate one or more steps are still required to create product of the pathway and these are not drawn explicitly. Note that this is also an example of pathway products feeding back to regulate upstream enzyme activity.
Attribution: Original work - Marc T. Facciotti