
S2019_Lecture_05_Reading
- Page ID
- 21450
Nucleic acids

There are two types of nucleic acids in biology: DNA and RNA. DNA carries the heritable genetic information of the cell and is composed of two antiparallel strands of nucleotides arranged in a helical structure. Each nucleotide subunit is composed of a pentose sugar (deoxyribose), a nitrogenous base, and a phosphate group. The two strands associate via hydrogen bonds between chemically complementary nitrogenous bases. Interactions known as "base stacking" interactions also help stabilize the double helix. By contrast to DNA, RNA can be either be single stranded, or double stranded. It too is composed of a pentose sugar (ribose), a nitrogenous base, and a phosphate group. RNA is a molecule of may tricks. It is involved in protein synthesis as a messenger, regulator, and catalyst of the process. RNA is also involved in various other cellular regulatory processes and helps to catalyze some key reactions (more on this later). With respect to RNA, in this course we are primarily interested in (a) knowing the basic molecular structure of RNA and what distinguishes it from DNA, (b) understanding the basic chemistry of RNA synthesis that occurs during a process called transcription, (c) appreciating the various roles that RNA can have in the cell, and (d) learning the major types of RNA that you will encounter most frequently (i.e. mRNA, rRNA, tRNA, miRNA etc.) and associating them with the processes they are involved with. In this module we focus primarily on the chemical structures of DNA and RNA and how they can be distinguished from one another.
Nucleotide structure
The two main types of nucleic acids are deoxyribonucleic acid (DNA) and ribonucleic acid (RNA). DNA and RNA are made up of monomers known as nucleotides. Individual nucleotides condense with one another to form a nucleic acid polymer. Each nucleotide is made up of three components: a nitrogenous base (for which there are five different types), a pentose sugar, and a phosphate group. These are depicted below. The main difference between these two types of nucleic acids is the presence or absence of a hydroxyl group at the C2 position, also called the 2' position (read "two prime"), of the pentose (see Figure 1 legend and section on the pentose sugar for more on carbon numbering). RNA has a hydroxyl functional group at that 2' position of the pentose sugar; the sugar is called ribose, hence the name ribonucleic acid. By contrast, DNA lacks the hydroxyl group at that position, hence the name, "deoxy" ribonucleic acid. DNA has a hydrogen atom at the 2' position.
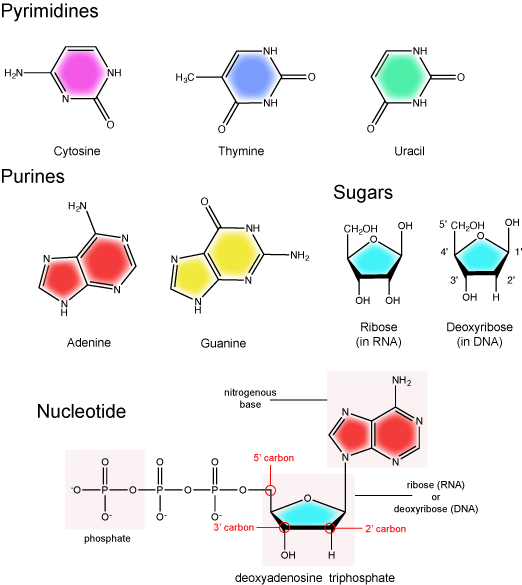
The nitrogenous base
The nitrogenous bases of nucleotides are organic molecules and are so named because they contain carbon and nitrogen. They are bases because they contain an amino group that has the potential of binding an extra hydrogen, and thus acting as a base by decreasing the hydrogen ion concentration in the local environment. Each nucleotide in DNA contains one of four possible nitrogenous bases: adenine (A), guanine (G), cytosine (C), and thymine (T). By contrast, RNA contains adenine (A), guanine (G) cytosine (C), and uracil (U) instead of thymine (T).
Adenine and guanine are classified as purines. The primary distinguishing structural feature of a purine is double carbon-nitrogen ring. Cytosine, thymine, and uracil are classified as pyrimidines. These are structurally distinguished by a single carbon-nitrogen ring. You will be expected to recognize that each of these ring structures is decorated by functional groups that may be involved in a variety of chemistries and interactions.
Note: practice
Take a moment to review the nitrogenous bases in Figure 1. Identify functional groups as described in class. For each functional group identified, describe what type of chemistry you expect it to be involved in. Try to identify whether the functional group can act as either a hydrogen bond donor, acceptor, or both?
The pentose sugar
The pentose sugar contains five carbon atoms. Each carbon atom of the sugar molecule are numbered as 1′, 2′, 3′, 4′, and 5′ (1′ is read as “one prime”). The two main functional groups that are attached to the sugar are often named in reference to the carbon to whch they are bound. For example, the phosphate residue is attached to the 5′ carbon of the sugar and the hydroxyl group is attached to the 3′ carbon of the sugar. We will often use the carbon number to refer to functional groups on nucleotides so be very familiar with the structure of the pentose sugar.
The pentose sugar in DNA is called deoxyribose, and in RNA, the sugar is ribose. The difference between the sugars is the presence of the hydroxyl group on the 2' carbon of the ribose and its absence on the 2' carbon of the deoxyribose. You can, therefore, determine if you are looking at a DNA or RNA nucleotide by the presence or absence of the hydroxyl group on the 2' carbon atom—you will likely be asked to do so on numerous occasions, including exams.
The phosphate group
There can be anywhere between one and three phosphate groups bound to the 5' carbon of the sugar. When one phosphate is bound, the nucleotide is referred to as a Nucleotide MonoPhosphate (NMP). If two phosphates are bound the nucleotide is referred to as Nucleotide DiPhosphate (NDP). When three phosphates are bound to the nucleotide it is referred to as a Nucleotide TriPhosphate (NTP). The phosphoanhydride bonds between that link the phosphate groups to each other have specific chemical properties that make them good for various biological functions. The hydrolysis of the bonds between the phosphate groups is thermodynamically exergonic in biological conditions; nature has evolved numerous mechanisms to couple this negative change in free energy to help drive many reactions in the cell. Figure 2 shows the structure of the nucleotide triphosphate Adenosine Triphosphate, ATP, that we will discuss in greater detail in other chapters.
Note: "high-energy" bonds
The term "high-energy bond" is used A LOT in biology. This term is, however, a verbal shortcuts that can cause some confusion. The term refers to the amount of negative free energy associated with the hydrolysis of the bond in question. The water (or other equivalent reaction partner) is an important contributor to the energy calculus. In ATP, for instance, simply "breaking" a phosphoanhydride bond - say with imaginary molecular tweezers - by pulling off a phosphate would not be energetically favorable. We must, therefore, be careful not to say that breaking bonds in ATP is energetically favorable or that it "releases energy". Rather, we should be more specific, noting that they hydrolysis of the bond is energetically favorable. Some of this common misconception is tied to, in our opinion, the use of the term "high energy bonds". While in Bis2a we have tried to minimize the use of the vernacular "high energy" when referring to bonds, trying instead to describe biochemical reactions by using more specific terms, as students of biology you will no doubt encounter the potentially misleading - though admittedly useful - short cut "high energy bond" as you continue in your studies. So, keep the above in mind when you are reading or listening to various discussions in biology. Heck, use the term yourself. Just make sure that you really understand what it refers to.

| Adenosine triphosphate |
Double helix structure of DNA
DNA has a double helix structure (shown below) created by two strands of covalently linked nucleotide subunits. The sugar and phosphate groups of each strand of nucleotides are positioned on the outside of the helix, forming the backbone of the DNA (highlighted by the orange ribbons in Figure 3). The two strands of the helix run in opposite directions, meaning that the 5′ carbon end of one strand will face the 3′ carbon end of its matching strand (See Figures 4 and 5). We referred to this orientation of the two strands as antiparallel. Note too that phosphate groups are depicted in Figure 3 as orange and red "sticks" protruding from the ribbon. The phosphates are negatively charged at physiological pHs and therefore give the backbone of the DNA a strong local negatively charged character. By contrast, the nitrogenous bases are stacked in the interior of the helix (these are depicted as green, blue, red, and white sticks in Figure 3). Pairs of nucleotides interact with one another through specific hydrogen bonds (shown in Figure 5). Each pair of separated from the next base pair in the ladder by 0.34 nm and this close stacking and planar orientation gives rise to energetically favorable base-stacking interactions. The specific chemistry associated with these interactions is beyond the content of Bis2a but is described in more detail here for the curious or more advanced students. We do expect, however, that students are aware that the stacking of the nitrogenous bases contributes to the stability of the double helix and defer to your upper-division genetics and organic chemistry instructors to fill in the chemical details.

In a double helix, certain combinations of base pairing are chemically more favored than others based on the types and locations of functional groups on the nitrogenous bases of each nucleotide. In biology we find that:
Adenine (A) is chemically complementary with thymidine (T) (A pairs with T)
and
Guanine (G) is chemically complementary with cytosine (C) (G pairs with C).
We often refer to this pattern as "base complementarity" and say that the antiparallel strands are complementary to each other. For example, if the sequence of one strand is of DNA is 5'-AATTGGCC-3', the complementary strand would have the sequence 5'-GGCCAATT-3'.
We sometimes choose to represent complementary double-helical structures in text by stacking the complementary strands on top of on another as follows:
5' - GGCCAATTCCATACTAGGT - 3'
3' - CCGGTTAAGGTATGATCCA - 5'
Note that each strand has its 5' and 3' ends labeled and that if one were to walk along each strand starting from the 5' end to the 3' end that the direction of travel would be opposite the other for each strand; the strands are antiparallel. We commonly say things like "running 5-prime to 3-prime" or "synthesized 5-prime to 3-prime" to refer to the direction we are reading a sequence or the direction of synthesis. Start getting yourself accustomed to this nomenclature.


Functions and roles of nucleotides and nucleic acids to look out for in Bis2a
In addition to their structural roles in DNA and RNA, nucleotides such as ATP and GTP also serve as mobile energy carriers for the cell. Some students are surprised when they learn to appreciate that the ATP and GTP molecules we discuss in the context of bioenergetics are the same as those involved in the formation of nucleic acids. We will cover this in more detail when we discuss DNA and RNA synthesis reactions. Nucleotides also play important roles as co-factors in many enzymatically catalyzed reactions.
Nucleic acids, RNA in particular, play a variety of roles in in cellular process besides being information storage molecules. Some of the roles that you should keep an eye out for as we progress through the course include: (a) Riboprotein complexes - RNA-Protein complexes in which the RNA serves both catalytic and structural roles. Examples of such complexes include, ribosomes (rRNA), RNases, splicesosome complexes, and telomerase. (b) Information storage and transfer roles. These roles include molecules like DNA, messenger RNA (mRNA), transfer RNA (tRNA). (c) Regulatory roles. Examples of these include various non-coding (ncRNA). Wikipedia has a comprehensive summary of the different types of known RNA molecules that we recommend browsing to get a better sense of the great functional diversity of these molecules.
Amino Acids

Amino acids are the monomers that make up proteins. Each amino acid has the same core structure, which consists of a central carbon atom, also known as the alpha (α) carbon, bonded to an amino group (NH2), a carboxyl group (COOH), and a hydrogen atom. Every amino acid also has another atom or group of atoms bonded to the alpha carbon known alternately as the R group, the variable group or the side-chain.

Amino acids have a central asymmetric carbon to which an amino group, a carboxyl group, a hydrogen atom, and a side chain (R group) are attached.
Attribution: Marc T. Facciotti (own work)
Note: Possible discussion
Recall that one of the learning goals for this class is that you (a) be able to recognize, in a molecular diagram, the backbone of an amino acid and its side chain (R-group) and (b) that you be able to draw a generic amino acid. Make sure that you practice both. You should be able to recreate something like the figure above from memory (a good use of your sketchbook is to practice drawing this structure until you can do it with the crutch of a book or the internet).
The Amino Acid Backbone
The name "amino acid" is derived from the fact that all amino acids contain both an amino group and carboxyl-acid-group in their backbone. There are 20 common amino acids present in natural proteins and each of these contain the same backbone. The backbone, when ignoring the hydrogen atoms, consists of the pattern:
N-C-C
When looking at a chain of amino acids it is always helpful to first orient yourself by finding this backbone pattern starting from the N terminus (the amino end of the first amino acid) to the C terminus (the carboxylic acid end of the last amino acid).
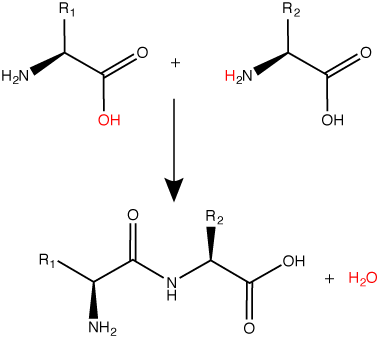
Peptide bond formation is a dehydration synthesis reaction. The carboxyl group of the first amino acid is linked to the amino group of the second incoming amino acid. In the process, a molecule of water is released and a peptide bond is formed.
Try finding the backbone in the dipeptide formed from this reaction. The pattern you are looking for is: N-C-C-N-C-C
Attribution: Marc T. Facciotti (own work)
The sequence and the number of amino acids ultimately determine the protein's shape, size, and function. Each amino acid is attached to another amino acid by a covalent bond, known as a peptide bond, which is formed by a dehydration synthesis (condensation) reaction. The carboxyl group of one amino acid and the amino group of the incoming amino acid combine, releasing a molecule of water and creating the peptide bond.
Amino Acid R group
The amino acid R group is a term that refers to the variable group on each amino acid. The amino acid backbone is identical on all amino acids, the R groups are different on all amino acids. For the structure of each amino acid refer to the figure below.

There are 20 common amino acids found in proteins, each with a different R group (variant group) that determines its chemical nature. R-groups are circled in teal. Charges are assigned assuming pH ~6.0. The full name, three letter abbreviation and single letter abbreviations are all shown.
Attribution: Marc T. Facciotti (own work)
| Glycine | Glutamate | Tryptophan |
Note: Possible Discussion
Let's think about the relevance of having 20 different amino acids. If you were using biology to build proteins from scratch, how might it be useful if you had 10 more different amino acids at your disposal? By the way, this is actually happening in a variety of research labs - why would this be potentially useful?
Each variable group on an amino acid gives that amino acid specific chemical properties (acidic, basic, polar, or nonpolar). You should be familiar with most of the functional groups in the R groups by now. The chemical properties associated with the whole collection of individual functional groups gives each amino acid R group unique chemical potential.
For example, amino acids such as valine, methionine, and alanine are typically nonpolar or hydrophobic in nature, while amino acids such as serine and threonine are said to have polar character and possess hydrophilic side chains.
Note: Practice
Using your knowledge of functional groups, try classifying each amino acid in the figure above as either having the tendency to be polar or nonpolar. Try to find other classification schemes and think make lists for yourself of the amino acids you would put into each group. You can also search the internet for amino acid classification schemes - you will notice that there are different ways of grouping these chemicals based on chemical properties. You may even find that there are contradictory schemes. Try to think about why this might be and apply your chemical logic to figuring out why certain classification schemes were adopted and why specific amino acids were placed in certain groups.
Tags recommended by the template: article:topic
Proteins

Proteins are class of biomolecules that perform a wide array of functions in biological systems. Some proteins serve as catalysts for specific biochemical reactions. Other proteins act as signaling molecules that allow cells to "talk" with one another. Proteins, like the keratin in fingernails, can also act in a structural capacity. While the variety of possible functions for proteins is remarkably diverse, all of these functions are encoded by a linear assembly of amino acids, each connected to their neighbor via a peptide bond. The unique composition (types of amino acids and the number of each) and the order in which they are linked together determine the final three dimensional form that the protein will adopt and therefore, also the protein's biological "function". Many proteins can, in a cellular environment, spontaneously and often rapidly take on their final form in a process called protein folding. To watch a short (four minutes) introduction video on protein structure click here.
Protein structure
Protein structures can be described by four different levels of structural organization called primary, secondary, tertiary, and quaternary structures. These are briefly introduced in the sections that follow.
Primary structure
The unique sequence of amino acids in a polypeptide chain is its primary structure (Figure 1). The amino acids in this chain are linked to one another other via a series of peptide bonds. The chain of amino acids is often referred to as a polypeptide (multiple peptides).
Due to the common backbone structure of amino acids, the resulting backbone of the protein has a repeating -N-Cα-C-N-Cα-C- pattern that can be readily identified in atomic resolution models of protein structures (Figure 2). Be aware that one of the learning goals for this class is for you to be able to examine a model like the one below and to identify the backbone from the side chain atoms (e.g. create the purple trace and blue shading if there aren't any). This can be done by finding the -N-Cα-C-N-Cα-C- pattern. Moreover, another learning goal for this class is that you are able to create drawings that model the structure of a typical protein backbone and its side chains (aka. variable group, R group). This task can be greatly simplified if you remember to start your model by first creating the -N-Cα-C-N-Cα-C- pattern and then filling in the variable groups.

Secondary structure
Due to the specific chemistry of the peptide bond the backbone between adjacent alpha-carbon atoms forms a highly planar structure (Figure 3). This means that all of the atoms linked by the pink quadrilateral lie on the same plane. The polypeptide is therefore structurally constrained since very little rotation can happen around the peptide bond itself. Rather, rotations occur around the two bonds extending away from the alpha carbons. These structural constraints lead to two commonly observed patterns of structure that are associated with the organization of the backbone itself.
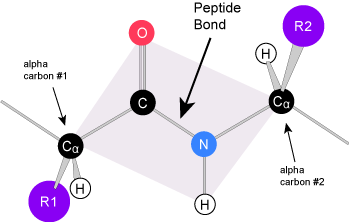
We call these patterns of backbone structure the secondary structure of the protein. The most common secondary structure patterns occurring via rotations of the bonds around each alpha-carbon, are the α-helix, β-sheet and loop structures. As the name suggests, the α-helix is characterized by a helical structure made by twisting the backbone. The β-sheet is actually the association between two or more structures called β-strands. If the orientation (N-terminus to C-terminus direction) of two associating β-strands are oriented in the same/parallel direction, the resulting β-sheet is called a parallel β-sheet. Meanwhile, if two associating β-strands are oriented in opposite/anti-parallel directions, the resulting β-sheet is called an anti-parallel β-sheet. The α-helix and β-sheet are both stabilized by hydrogen bonds that form between backbones atoms of amino acids in close proximity to one another. More specifically, the oxygen atom in the carbonyl group from one amino acid can form a hydrogen bond with a hydrogen atom bound to the nitrogen in the amino group of another amino acid. Loop structures by contrast refer to all secondary structure (e.g. backbone structure) that can not be identified as either α-helix or β-sheet.

Tertiary structure
The backbone and secondary structure elements will further fold into a unique and relatively stable three-dimensional structure called the tertiary structure of the protein. The tertiary structure is what we typically associate with the "functional" form of a protein. In Figure 6 two examples of tertiary structure are shown. In both structures, the protein is abstracted into a "cartoon" that depicts the polypeptide chain as a single continuous line or ribbon tracing the path between alpha carbons of amino acids linked to one another by peptide bonds - the ribbon traces the backbone of the protein (Figure 5).

The ribbon created by joining alpha-carbons can be drawn as a simple continuous line or it can be enhanced by uniquely representing secondary structural elements. For instance, when an α-helix is identified, the helix is usually highlighted by accentuating/broadening the ribbon to make the helical structure stand out. When a β-strand is present, the ribbon is usually broadened and an arrow is typically added to the C-terminal end of each β-strand - the arrow helps to identify the orientation of the polypeptide and whether β-sheets are parallel or anti-parallel. The thin ribbon connecting α-helix and β-strand elements is used to represent the loops. Loops in proteins can be highly structured and play an important role in the protein's function. They should not be treated lightly or dismissed as unimportant because their name lacks a Greek letter.

| Crystallin (PDBID 1a45) | Triose phosphate isomerase (PDBID 1tim) |
The tertiary structure is the product of many different types of chemical interactions among amino acid R groups, backbone atoms, ions in solution and water. These bonds include ionic, covalent, and hydrogen bonds and Van der Waals interactions. For example, ionic bonds may form between various ionizable side chains. It may, for instance, be energetically favorable for a negatively charged R group (e.g. an Aspartate) to interact with a positively charged R group (e.g. an Arginine). The resulting ionic interaction may then become part of the network of interactions that helps to stabilize the three dimensional fold of the protein. By contrast, R groups with like charges will likely be repelled by each other and be therefore unlikely to form a stable association thereby disfavoring a structure that would include that association. Likewise, hydrogen bonds may form between various R groups or between R groups and backbone atoms. These hydrogen bonds may also contribute to stabilizing the tertiary structure of the protein. In some cases covalent bonds may also form between amino acids. The most commonly observed covalent linkage between amino acids involves two cysteines and is termed a disulfide bond or disulfide linkage.
Finally, the association of the protein's functional groups with water also helps to drive chemical associations that help to stabilize the final protein structure. The interactions with water can, of course, include the formation of hydrogen bonds between polar functional groups on the protein and water molecules. Perhaps more importantly, however, is the drive for the protein to avoid placing too many hydrophobic functional groups in contact with water. The result of this desire to avoid interactions between water and hydrophobic functional groups means that the less polar side chains will often associate with one another away from water resulting in some energetically favorable Van der Waals interactions and the avoidance of energetic penalties associated with exposing the non-polar side chains to water. Indeed, the energetic penalty is so high for "exposing" the non-polar side chains to water that burying these groups away from water is thought to be one of the primary energetic drivers of protein folding and stabilizing forces holding the protein together in its tertiary structure.
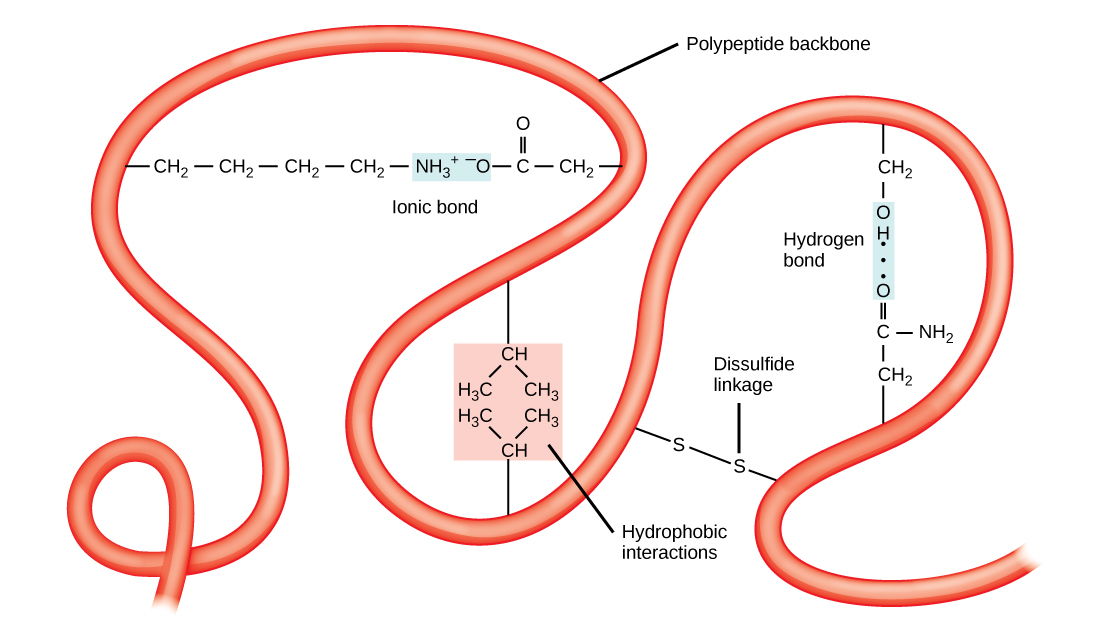
Quaternary structure
In nature, the functional forms of some proteins are formed by the close association of several polypeptides. In such cases the individual polypeptides are also known as subunits. When the functional form of a protein requires the assembly of two or more subunits we call this level of structural organization the protein's quaternary structure. Yet again, combinations of ionic, hydrogen, and covalent bonds together with Van der Waals associations that occur through the "burial" of hydrophobic group at the interfaces between subunits help to stabilize the quaternary structures of proteins.

Denaturation
As was previously described, each protein has its own unique structure that is held together by various types of chemical interactions. If the protein is subject to changes in temperature, pH, or exposure to chemicals, that change the nature of or interfere with the associations between functional groups, the protein's secondary, tertiary and/or quaternary structures may change, even though the primary structure remains the same. This process is known as denaturation. While in the test tube denaturation is often reversible, in the cell the process can often be, for practical purposes, irreversible, leading to loss of function and the eventual recycling of the protein's amino acids. Resistance to environmental stresses that can lead to denaturation vary greatly amongst the proteins found in nature. For instance, some proteins are remarkably resistant to high temperatures; for instance, bacteria that survive in hot springs have proteins that function at temperatures close to the boiling point of water. Some proteins are able to withstand the very acidic, low pH, environment of the stomach. Meanwhile some proteins are very sensitive to organic solvents while others can be found that are remarkably tolerant of these chemicals (the latter are prized for use in various industrial processes).
Finally, while many proteins can form their three dimensional structures completely on their own, in many cases proteins often receive assistance in the folding process from protein helpers known as chaperones (or chaperonins) that associate with their protein targets during the folding process. The chaperones are thought to act by minimizing the aggregation of polypeptides into non-functional forms - a process that can occur through the formation of non-ideal chemical associations.