Generation of NADPH and ATP
The overall function of light-dependent reactions is to transfer solar energy into chemical compounds, largely the molecules NADPH and ATP. This energy supports the light-independent reactions and fuels the assembly of sugar molecules. We depict the light-dependent reactions in Figures 6 and 7. Protein complexes and pigment molecules work together to produce NADPH and ATP.

Figure 1. A photosystem consists of a light-harvesting complex and a reaction center. Pigments in the light-harvesting complex pass light energy to two special chlorophyll a molecules in the reaction center. The light excites an electron from the chlorophyll a pair, which passes to the primary electron acceptor. The excited electron must then be replaced. In (a) photosystem II, the electron comes from the splitting of water, which releases oxygen as a waste product. In (b) photosystem I, the electron comes from the chloroplast electron transport chain discussed below.
The actual step that transfers light energy into a biomolecule takes place in a multiprotein complex called a photosystem, two types of which are found embedded in the thylakoid membrane, photosystem II (PSII) and photosystem I (PSI). The two complexes differ based on what they oxidize (that is, the source of the low energy electron supply) and what they reduce (the place to which they deliver their energized electrons).
Both photosystems have the same basic structure; several antenna proteins to which the chlorophyll molecules bind surround the reaction center in which the photochemistry takes place. Each photosystem associates with the light-harvesting complex, which passes energy captured from sunlight to the reaction center; it comprises multiple antenna proteins that contain a mixture of 300–400 chlorophyll a and b molecules and other pigments like carotenoids. The absorption of a single photon—a distinct quantity or “packet” of light—by any of the chlorophylls pushes that molecule into an excited state. In short, the biological molecule has now captured light energy. However, the energy is not yet stored in any useful form. The captured energy transfers from chlorophyll to chlorophyll until, eventually (after about a millionth of a second), it arrives to the reaction center. Up to this point, only energy has been transferred between molecules, not electrons.

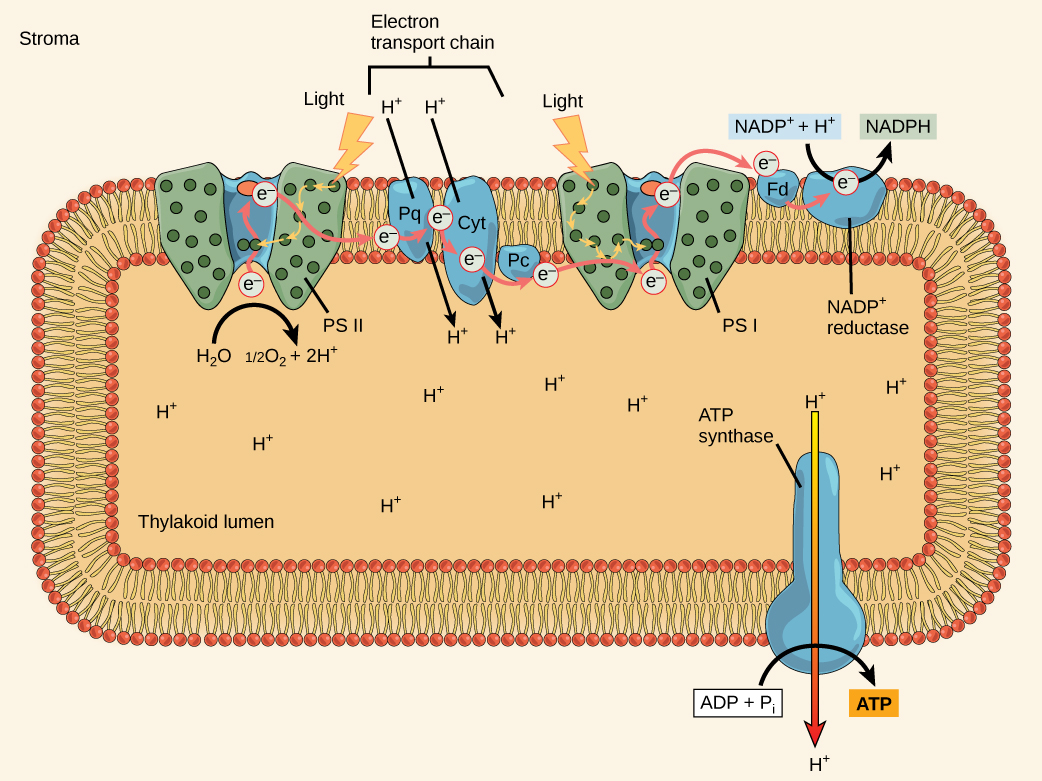
Figure 2. In the photosystem II (PSII) reaction center, energy from sunlight is used to extract electrons from water. The electrons travel through the chloroplast electron transport chain to photosystem I (PSI), which reduces NADP+ to NADPH. The electron transport chain moves protons across the thylakoid membrane into the lumen. At the same time, splitting of water adds protons to the lumen, and reduction of NADPH removes protons from the stroma. The net result is a low pH in the thylakoid lumen, and a high pH in the stroma. ATP synthase uses this electrochemical gradient to make ATP.
The reaction center contains a pair of chlorophyll a molecules with a special property. Those two chlorophylls can undergo oxidation upon excitation; they can give up an electron in a process called a photoactivation. It is at this step in the reaction center, this step in photophosphorylation, that light energy transfers into an excited electron. All the subsequent steps involve a getting of that electron onto the energy carrier NADPH for delivery to the Calvin cycle where the electron can be deposited onto carbon for long-term storage in the form of a carbohydrate.
The Z-scheme
PSII and PSI are two major components of the photosynthetic electron transport chain, which also include the cytochrome complex. The reaction center of PSII (called P680) delivers its high-energy electrons, one at a time, to a primary electron acceptor called pheophytin (Ph), and then sequentially to two bound plastoquinones QA and QB. Electrons then transfer off of PSII onto a pool of mobile plastoquinones (Q pool) which then transfer the electrons to a protein complex called Cytochromeb6f. The cytochrome complex uses the red/ox transfers to pump proton across the thylakoyd membrane establishing a proton-motive force that can be used for the synthesis of ATP. Electrons leaving the Cytochrome transfer to a copper-containing protein called plastocyanin (PC) which then transfers electrons to PSI (P700). P680’s missing electron is replaced by extracting an electron from water; thus, water splits and PSII is re-reduced after every photoactivation step. Just for the sake of sharing some numbers: Splitting one H2O molecule releases two electrons, two hydrogen atoms, and one atom of oxygen. The formation of one molecule of diatomic O2 gas requires the splitting two water molecules. In plant tissue, mitochondria use about ten percent of that oxygen to support oxidative phosphorylation. The rest escapes to the atmosphere where it is used by aerobic organisms to support respiration.
As electrons move through the proteins that live between PSII and PSI, they take part in exergonic red/ox transfers. The free energy associated with the exergonic red/ox reaction is coupled to the endergonic transport of protons from the stromal side of the membrane to the thylakoid lumen by the cytochrome complex. Those hydrogen ions, plus the ones produced by splitting water, accumulate in the thylakoid lumen and create a proton motive force that will drive the synthesis of ATP in a later step. Since the electrons on PSI now have a greater reduction potential than when they started their trek (it is important to note that unexcited PSI has a greater red/ox potential than NADP+/NADPH), they must be re-energized in PSI before getting deposited onto NADP+. Therefore, to complete this process, another photon must be absorbed by the PSI antenna. That energy transfers to the PSI reaction center (called P700). P700 then oxidizes and sends an electron through several intermediate red/ox steps to NADP+ to form NADPH. Thus, PSII captures the energy in light and couples its transfer via red/ox reactions to the creation of a proton gradient. As already noted, the exergonic and controlled relaxation of this gradient can be coupled to the synthesis of ATP. PSI captures energy in light and couples that, through a series of red/ox reactions, to reduce NADP+ into NADPH. The two photosystems work in concert, in part, to guarantee that the production of NADPH will be in the right proportion to the production of ATP. Other mechanisms exist to fine tune that ratio to match the chloroplast’s constantly changing energy needs.

Figure 3. A diagram depicting the flow of electrons and the red/ox potentials of their carriers in oxygenic photosynthetic systems expressing both photosystem I (boxed in blue) and photosystem II (boxed in green). Ph = pheophytin; QA = bound plastoquinone, QB = more loosely associated plastoquinone; Q pool = mobile plastoquinone pool; Cyt bf = Cytochrome b6f complex; PC = plastocyanin; Chla0 = special chrolophyl; A1 = vitamin K; Fx and FAB = iron-sulfur centers; Fd = ferredoxin; FNR = ferredoxin-NADP reductase. Attribution: Marc T. Facciotti (own work)