Transport across the membrane
Design challenge problem and subproblems
General Problem: The cell membrane must simultaneously act as a barrier between "IN" and "OUT" and control specifically which substances enter and leave the cell and how quickly and efficiently they do so.
Subproblems: The chemical properties of molecules that must enter and leave the cell are highly variable. Some subproblems associated with this are: (a) Large and small molecules or collections of molecules must be able to pass across the membrane. (b) Both hydrophobic and hydrophilic substances must have access to transport. (c) Substances must be able to cross the membrane with and against concentration gradients. (d) Some molecules look very similar (e.g. Na+ and K+) but transport mechanisms must still be able to distinguish between them.
Energy story perspective
We can consider transport across a membrane from an energy story perspective; it is a process after all. For instance, at the beginning of the process a generic substance X may be on the inside or outside of the cell. At the end of the process, the substance will be on the opposite side from which it started.
e.g. X(in) ---> X(out),
where in and out refer to inside the cell and outside the cell, respectively.
At the beginning the matter in the system might be a very complicated collection of molecules inside and outside of the cell but with one molecule of X more inside the cell than out. At the end, there is one more molecule of X on the outside of the cell and one less on the inside. The energy in the system at the beginning is stored largely in the molecular structures and their motions and in electrical and chemical concentration imbalances across the cell membrane. The transport of X out of the cell will not change the energies of the molecular structures significantly but it will change the energy associated with the imbalance of concentration and or charge across the membrane. That is the transport will, like all other reactions, be exergonic or endergonic. Finally, some mechanism or sets of mechanisms of transport will need to be described.
Selective permeability
One of the great wonders of the cell membrane is its ability to regulate the concentration of substances inside the cell. These substances include: ions such as Ca2+, Na+, K+, and Cl–; nutrients including sugars, fatty acids, and amino acids; and waste products, particularly carbon dioxide (CO2), which must leave the cell.
The membrane’s lipid bilayer structure provides the first level of control. The phospholipids pack tightly, and the membrane has a hydrophobic interior. This structure alone creates what is known as a selectively permeable barrier, one that only allows substances meeting certain physical criteria to pass through it. In the case of the cell membrane, only relatively small, nonpolar materials can move through the lipid bilayer at biologically relevant rates (remember, the lipid tails of the membrane are nonpolar).
Selective permeability of the cell membrane refers to its ability to differentiate between different molecules, only allowing some molecules through while blocking others. Some of this selective property stems from the intrinsic diffusion rates for different molecules across a membrane. A second factor affecting the relative rates of movement of various substances across a biological membrane is the activity of various protein-based membrane transporters, both passive and active, that will be discussed in more detail in subsequent sections. First, we take on the notion of intrinsic rates of diffusion across the membrane.
Relative permeability
That different substances might cross a biological membrane at different rates should be relatively intuitive. There are differences in the mosaic composition of membranes in biology and differences in the sizes, flexibility, and chemical properties of molecules so it stands to reason that the permeability rates vary. It is a complicated landscape. The permeability of a substance across a biological membrane can be measured experimentally and we can report the rate of movement across a membrane in what are known as membrane permeability coefficients.
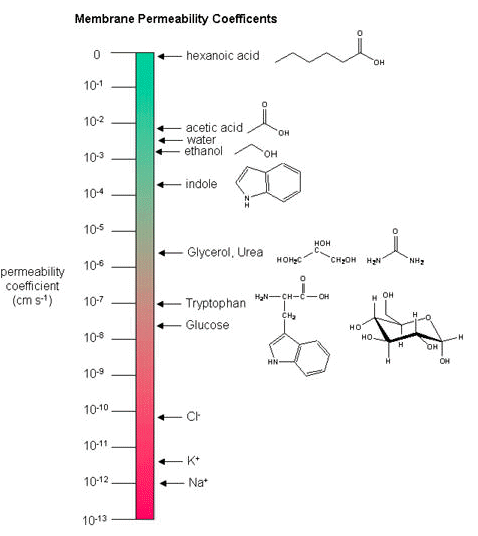
Membrane permeability coefficients
Below, we plot a variety of compounds with respect to their membrane permeability coefficients (MPC) as measured against a simple biochemical approximation of a real biological membrane. The reported permeability coefficient for this system is the rate at which simple diffusion through a membrane occurs and is reported in units of centimeters per second (cm/s). The permeability coefficient is proportional to the partition coefficient and is inversely proportional to the membrane thickness.
It is important that you can read and interpret the diagram below. The larger the coefficient, the more permeable the membrane is to the solute. For example, hexanoic acid is very permeable, a MPC of 0.9; acetic acid, water, and ethanol have MPCs between 0.01 and 0.001, and they are less permeable than hexanoic acid. Whereas ions, such as sodium (Na+), have an MPC of 10-12, and cross the membrane at a comparatively slow rate.
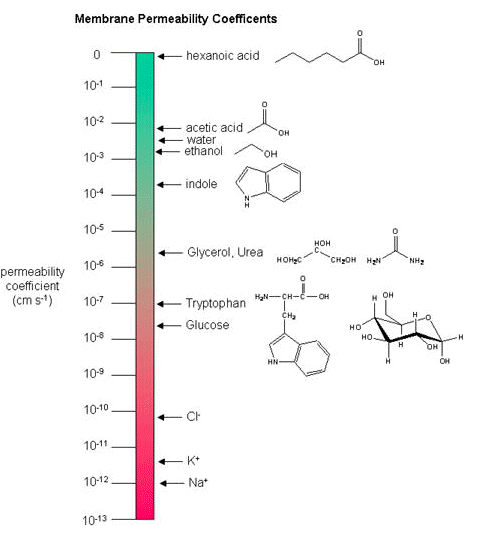
While there are certain trends or chemical properties that can be roughly associated with a different compound permeability (small things go through "fast", big things "slowly", charged things not at all etc.), we caution against over-generalizing. The molecular determinants of membrane permeability are complicated and involve many factors including: the specific composition of the membrane, temperature, ionic composition, hydration; the chemical properties of the solute; the potential chemical interactions between the solute in solution and in the membrane; the dielectric properties of materials; and the energy trade-offs associated with moving substances into and out of various environments. So, in this class, rather than try to apply "rules" and try to develop too many arbitrary "cut-offs", we will strive to develop a general sense of some properties that can influence permeability and leave the assignment of absolute permeability to experimentally reported rates. In addition, we will also try to minimize the use of vocabulary that depends on a frame of reference. For instance, saying that compound A diffuses "quickly" or "slowly" across a bilayer only means something if the terms "quickly" or "slow" are numerically defined or the biological context is understood.


 Selective permeability
Selective permeability