
8.8: Reproductive Strategies
- Page ID
- 75006
Population ecologists have hypothesized that suites of characteristics may evolve in species that lead to particular adaptations to their environments. These adaptations impact the kind of population growth their species experience. Population ecologists have described a continuum of life-history “strategies” with K-selected species on one end and r-selected species on the other (table \(\PageIndex{a}\)). K-selected species are adapted to stable, predictable environments. Populations of K-selected species tend to exist close to their carrying capacity (which is represented by the letter "K" in the equation for logistic population growth). These species tend to have larger, but fewer, offspring, contribute large amounts of resources to each offspring, and have long generation times. Elephants would be an example of a K-selected species (figure \(\PageIndex{a}\)). When a habitat becomes filled with a diverse collection of creatures competing with one another for the necessities of life, the advantage shifts to K-strategists. K-strategists have stable populations that are close to K. There is nothing to be gained from a high growth rate (r). The species will benefit most by a close adaptation to the conditions of its environment.
| Characteristics of K-selected species | Characteristics of r-selected species |
|---|---|
| Mature late | Mature early |
| Greater longevity | Lower longevity |
| Increased parental care | Decreased parental care |
| Strong competitors | Weak competitors (strong colonizers) |
| Fewer offspring | More offspring |
| Larger offspring | Smaller offspring |

r-selected species are adapted to unstable and unpredictable environments. They have large numbers of small offspring. Animals that are r-selected do not provide a lot of resources or parental care to offspring, and the offspring are relatively self-sufficient at birth. Examples of r-selected species are marine invertebrates such as jellyfish and plants such as the dandelion (figure \(\PageIndex{a}\)). r-strategists have short life spans and reproduce quickly, resulting in short generation times. For example, the housefly can produce 7 generations each year (each of about 120 young). Ragweed is well-adapted to exploiting its environment in a hurry - before competitors can become established. It grows rapidly and produces a huge number of seeds (after releasing its pollen, the bane of many hay fever sufferers). Because ragweed's approach to continued survival is through rapid reproduction (a high value of r) it is called an r-strategist. Other weeds, many insects, and many rodents are also r-strategists. If fact, if we consider an organism a pest, it is probably an r-strategist.
The two extreme strategies are at two ends of a continuum on which real species life histories will exist. In addition, life history strategies do not need to evolve as suites but can evolve independently of each other; therefore, each species may have some characteristics that trend toward one extreme or the other. Nevertheless, the r- and K-selection theory provides a foundation for a more accurate life history framework. New demographic-based models of life history evolution have been developed, which incorporate many ecological concepts included in r- and K-selection theory as well as population age structure and mortality factors.
Another tool used by population ecologists is a survivorship curve, which is a graph of the number of individuals surviving at each age interval versus time. These curves allow us to compare the life histories of different populations (figure \(\PageIndex{c}\)). There are three types of survivorship curves. In a type I survivorship curve, mortality is low in the early and middle years and occurs mostly in older individuals. Organisms exhibiting a type I survivorship typically produce few offspring and provide good care to the offspring increasing the likelihood of their survival. Humans and most mammals exhibit a type I survivorship curve. In type II survivorship curves, mortality is relatively constant throughout the entire life span, and mortality is equally likely to occur at any point in the life span. Many bird populations provide examples of an intermediate or type II survivorship curve. In type III survivorship curves, early ages experience the highest mortality with much lower mortality rates for organisms that make it to advanced years. Type III organisms typically produce large numbers of offspring, but provide very little or no care for them. Trees and marine invertebrates exhibit a type III survivorship curve because very few of these organisms survive their younger years, but those that do make it to an old age are more likely to survive for a relatively long period of time.
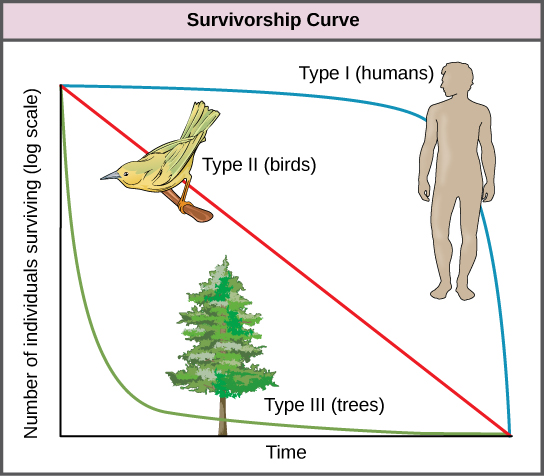
Figure \(\PageIndex{c}\): Survivorship curves show the chance of survival in a population according to age. The number of individuals surviving (on a log scale) is on the y-axis, and time (representing relative age) is on the x-axis. Humans and most mammals have a Type I survivorship curve, because death primarily occurs in the older years. The Type I survivorship curve is represented by a convex curve, which rapidly declines at the end. Birds have a Type II survivorship curve, as death at any age is equally probable. The Type II survivorship curve is represented by a straight line, going from the upper left to lower right of the graph. Trees have a Type III survivorship curve because very few survive the younger years, but after a certain age, individuals are much more likely to survive. The Type III survivorship curve is represented by a concave curve, which initially declines rapidly and then flattens.
While is no exact association between reproductive strategies (K- or r-selected) and survivorship curves (Type I, II, or III), K-selected species are more likely to have a Type III survivorship curve. r-selected species tend to have a Type I survivorship curve.
Attribution
Modified by Melissa Ha from the following sources:
- Population Dynamics and Regulation from General Biology by OpenStax (CC-BY)
- Population Demographics and Dynamics from Environmental Biology by Matthew R. Fisher (CC-BY)
- Principles of Population Growth from Biology by John W. Kimball (CC-BY)