
14.2: Genes and Chromatin in Eukaryotes
- Page ID
- 42834

Chromosomes and chromatin are a uniquely eukaryotic association of DNA with more or less protein. Bacterial DNA (and prokaryotic DNA generally) is relatively ‘naked’ – not visibly associated with protein.
The electron micrograph of an interphase cell (below) reveals that the chromatin can itself exist in various states of condensation.

Chromatin is maximally condensed during mitosis, forming chromosomes. During interphase, chromatin exists in more or less condensed forms, called Heterochromatin and euchromatin respectively. Transition between these chromatin forms involve changes in the amounts and types of proteins bound to the chromatin, and can that can occur during gene regulation, i.e., when genes are turned on or off. Active genes tend to be in the more dispersed euchromatin so that enzymes of replication and transcription have easier access to the DNA. Genes that are inactive in transcription are heterochromatic, obscured by additional chromatin proteins present in heterochromatin. We’ll be looking at some experiments that demonstrate this in a later chapter.
We can define three levels of chromatin organization in general terms:
1. DNA wrapped around histone proteins form nucleosomes in a "beads on a string" structure.
2. Multiple nucleosomes coil (condense), forming 30 nm fiber (solenoid) structures.
3. Higher-order packing of the 30 nm fiber leads to formation of metaphase chromosomes seen in mitosis & meiosis.
The levels of chromatin structure were determined in part by selective isolation and extraction of interphase cell chromatin, followed by selective chemical extraction of chromatin components. The steps are:
· Nuclei are first isolated from the cells.
· The nuclear envelope gently ruptured so as not to physically disrupt chromatin structure.
· the chromatin can be gently extracted by one of several different chemical treatments (high salt, low salt, acid...).
The levels of chromatin structure are illustrated below.

Salt extraction dissociates most of the proteins from the chromatin. When a low salt extract is centrifuged and the pellet resuspended, the remaining chromatin looks like beads on a string. DNA-wrapped nucleosomes are the beads, which are in turn linked by uniform lengths of metaphorical DNA “string’ ( # 1 in the illustration above). A high salt chromatin extract appears as a coil of nucleosomes, or 30 nm solenoid fiber (# 2 above). Other extraction protocols revealed other aspects of chromatin structure shown in #s 3 and 4 above. Chromosomes seen in metaphase of mitosis are the ‘highest order’, most condensed form of chromatin.
The 10 nm filament of nucleosome ‘beads-on-a-string’ remaining after a low salt extraction can be seen in an electron microscope as shown below.

When these nucleosome necklaces were digested with the enzyme deoxyribonuclease (DNAse), the DNA between the ‘beads’ was degraded, leaving behind shortened 10nm filaments after a short digest period, or just single beads the beads after a longer digestion (below).

Roger Kornberg (son of Nobel Laureate Arthur Kornberg who discovered the first DNA polymerase enzyme of replication) participated in the discovery and characterization of nucleosomes while he was still a post-doc! Electrophoresis of DNA extracted from these digests revealed nucleosomes separated by a “linker” DNA stretch of about 80 base pairs. DNA extracted from the nucleosomes was about 147 base pairs long. This is the DNA that had been wrapped around the proteins of the nucleosome.
After separating all of the proteins from nucleosomal DNA, five proteins were identified (illustrated below).
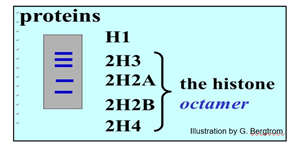
Histones are basic proteins containing many lysine and arginine amino acids. Their positively charged side chains enable these amino acids bind the acidic, negatively charged phosphodiester backbone of double helical DNA. The DNA wraps around an octamer of histones (2 each of 4 of the histone proteins) to form the nucleosome. About a gram of histones is associated with each gram of DNA. After a high salt chromatin extraction, the structure visible in the electron microscope is the 30nm solenoid, the coil of nucleosomes modeled in the figure below.

As shown above, simply increasing the salt concentration of an already extracted nucleosome preparation will cause the ‘necklace’ to fold into the 30nm solenoid structure.
173 Chromatin Structure: Dissecting Chromatin
As you might guess, an acidic extraction of chromatin should selectively remove the basic histone proteins, leaving behind an association of DNA with non-histone proteins. This proved to be the case. An electron micrograph of the chromatin remnant after an acid extraction of metaphase chromosomes is shown on the next page.
DNA freed of the regularly spaced histone-based nucleosomes, loops out, away from the long axis of the chromatin. Dark material along this axis is a protein scaffolding that makes up what’s left after histone extraction. Much of this protein is topoisomerase, an enzyme that prevents DNA from breaking apart under the strain of replication.
