
10.1: DNA is the Genetic Material
- Page ID
- 42814

By the early 1900’s, biochemists had isolated hundreds of different chemicals from living cells. Which of these was the genetic material? Proteins seemed like promising candidates, since they were abundant, diverse, and complex molecules. However, a few key experiments demonstrated that DNA, rather than protein, is the genetic material.
Griffith’s Transformation Experiment (1928)
Microbiologists identified two strains of the bacterium Streptococcus pneumoniae. The R-strain produced rough colonies on a bacterial plate, while the other S-strain was smooth (Figure \(\PageIndex{2}\)). More importantly, the S-strain bacteria caused fatal infections when injected into mice, while the R-strain did not (top, Figure \(\PageIndex{3}\)). Neither did “heat-treated” S-strain cells. Griffith in 1929 noticed that upon mixing “heat-treated” S-strain cells together with some R-type bacteria (neither should kill the mice), the mice died and there were S-strain, pathogenic cells recoverable. Thus, some non-living component from the S-type strains contained genetic information that could be transferred to and transform the living R-type strain cells into S-type cells.



Avery, MacLeod and McCarty’s Experiment (1944)
What kind of molecule from within the S-type cells was responsible for the transformation? To answer this, researchers named Avery, MacLeod and McCarty separated the S-type cells into various components, such as proteins, polysaccharides, lipids, and nucleic acids. Only the nucleic acids from S-type cells were able to make the R-strains smooth and fatal. Furthermore, when cellular extracts of S-type cells were treated with DNase (an enzyme that digests DNA), the transformation ability was lost. The researchers therefore concluded that DNA was the genetic material, which in this case controlled the appearance (smooth or rough) and pathogenicity of the bacteria.
Hershey and Chase’s Experiment (1952)
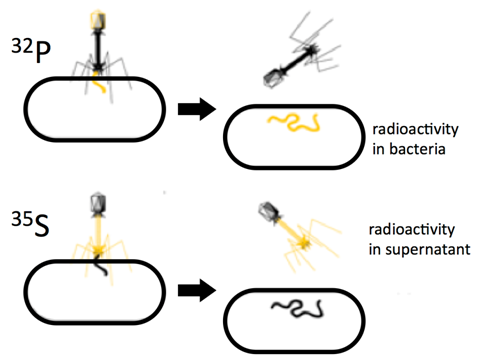
Further evidence that DNA is the genetic material came from experiments conducted by Hershey and Chase. These researchers studied the transmission of genetic information in a virus called the T2 bacteriophage, which used Escherichia coli as its host bacterium (Figure \(\PageIndex{4}\)). Like all viruses, T2 hijacks the cellular machinery of its host to manufacture more viruses. The T2 phage itself only contains both protein and DNA, but no other class of potential genetic material.

To determine which of these two types of molecules contained the genetic blueprint for the virus, Hershey and Chase grew viral cultures in the presence of radioactive isotopes of either phosphorus (32P) or sulphur (35S). The phage incorporated these isotopes into their DNA and proteins, respectively (Fig 1.5). The researchers then infected E. coli with the radiolabeled viruses, and looked to see whether 32P or 35S entered the bacteria. After ensuring that all viruses had been removed from the surface of the cells, the researchers observed that infection with 32P labeled viruses (but not the 35S labeled viruses) resulted in radioactive bacteria. This demonstrated that DNA was the material that contained genetic instructions.
Meselson and Stahl experiment (1958)
From the complementary strands model of DNA, proposed by Watson and Crick in 1953, there were three straightforward possible mechanisms for DNA replication: (1) semi-conservative, (2) conservative, and (3) dispersive (Fig 1.6).
- The semi-conservative model proposes the two strands of a DNA molecule separate during replication and then strand acts as a template for synthesis of a new, complementary strand.
- The conservative model proposes that the entire DNA duplex acts as a single template for the synthesis of an entirely new duplex.
- The dispersive model has the two strands of the double helix breaking into units that which are then replicated and reassembled, with the new duplexes containing alternating segments from one strand to the other.

Each of these three models makes a different prediction about the how DNA strands should be distributed following two rounds of replication. These predictions can be tested in the following experiment by following the nitrogen component in DNA in E. coli as it goes through several rounds of replication. Meselson and Stahl used different isotopes of Nitrogen, which is a major component in DNA. Nitrogen-14 (14N) is the most abundant natural isotope, while Nitrogen-15 (15N) is rare, but also denser. Neither is radioactive; each can be followed by a difference in density – “light” 14 vs “heavy”15 atomic weight in a CsCl density gradient ultra-centrifugation of DNA.
The experiment starts with E. coli grown for several generations on medium containing only 15N. It will have denser DNA. When extracted and separated in a CsCl density gradient tube, this “heavy” DNA will move to a position nearer the bottom of the tube in the more dense solution of CsCl (left side in Figure \(\PageIndex{7}\)). DNA extracted from E. coli grown on normal, 14 N containing medium will migrate more towards the less dense top of the tube.
If these E. coli cells are transferred to a medium containing only 14N, the “light” isotope, and grown for one generation, then their DNA will be composed of one-half 15N and one-half 14N. If the this DNA is extracted and applied to a CsCl gradient, the observed result is that one band appears at the point midway between the locations predicted for wholly 15N DNA and wholly 14N DNA (Figure \(\PageIndex{7}\)). This “single-band” observation is inconsistent with the predicted outcome from the conservative model of DNA replication (disproves this model), but is consistent with both that expected for the semi-conservative and dispersive models.
If the E. coli is permitted to go through another round of replication in the 14N medium, and the DNA extracted and separated on a CsCl gradient tube, then two bands were seen by Meselson and Shahl: one at the 14N-15N intermediate position and one at the wholly 14N position (Figure \(\PageIndex{7}\)). This result is inconsistent with the dispersive model (a single band between the 14N-15N position and the wholly 14 N position) and thus disproves this model. The two band observation is consistent with the semi-conservative model which predicts one wholly 14 N duplex and one 14N-15N duplex. Additional rounds of replication also support the semi-conservative model/hypothesis of DNA replication. Thus, the semi-conservative model is the currently accepted mechanism for DNA replication. Note however, that we now also know from more recent experiments that whole chromosomes, which can be millions of bases in length, are also semi-conservatively replicated.

These experiments, published in 1958, are a wonderful example of how science works. Researchers start with three clearly defined models (hypotheses). These models were tested, and two (conservative and dispersive) were found to be inconsistent with the observations and thus disproven. The third hypothesis, semi-conservative, was consistent with the observations and thereby supported and accepted as mechanism of DNA replication. Note, however, this is not “proof” of the model, just strong evidence for it; hypotheses are not “proven”, only disproven or supported.
RNA and protein
While DNA is the genetic material for the vast majority of organisms, there are some viruses that use RNA as their genetic material. These viruses can be either single or double stranded and include SARS, influenza, hepatitis C and polio, as well as the retroviruses like HIV-AIDS. Typically there is DNA used at some stage in their life cycle to replicate their RNA genome.
Also, the case of Prion infections agents transmit characteristics via only a protein (no nucleic acid present). Prions infect by transmitting a misfolded protein state from one aberrant protein molecule to a normally folded molecule. These agents are responsible for bovine spongiform encephalopathy (BSE, also known as "mad cow disease") in cattle and deer and Creutzfeldt–Jakob disease (CJD) in humans. All known prion diseases act by altering the structure of the brain or other neural tissue and all are currently untreatable and ultimately fatal.