
7.4: Mutations and Cancer
- Page ID
- 22487

Your cells may grow and divide without performing their necessary functions, or without fully replicating their DNA, or without copying their organelles. Probably not much good could come of that. So the cell cycle needs to be highly regulated and tightly controlled. And it is.
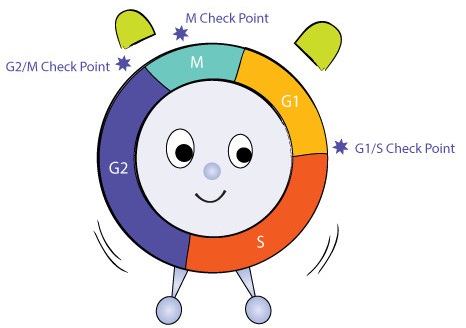
Control of the Cell Cycle
How does the cell know when to divide? How does the cell know when to replicate its DNA? How does the cell know when to proceed into mitosis or cytokinesis? The answers to these questions have to do with the control of the cell cycle. But how is the cell cycle controlled or regulated? Regulation of the cell cycle involves processes crucial to the survival of a cell. These include the detection and repair of damage to DNA, as well as the prevention of uncontrolled cell division. Uncontrolled cell division can be deadly to an organism; its prevention is critical for survival.
Cyclins and Kinases
The cell cycle is controlled by a number of protein-controlled feedback processes. Two types of proteins involved in the control of the cell cycle are kinases and cyclins. Cyclins activate kinases by binding to them, specifically they activate cyclin-dependent kinases (CDK). Kinases are enzymes that catalyze the transfer of a phosphate group from ATP to another molecule in a cell. They function as a control switch in many cellular functions, turning a function on or off, and regulating other cellular processes. Many times they are involved in activating a cascade of reactions. Cyclins comprise a group of proteins that are rapidly produced at key stages in the cell cycle. Once activated by a cyclin, CDK enzymes activate or inactivate other target molecules through phosphorylation. It is this precise regulation of proteins that triggers advancement through the cell cycle. Leland H. Hartwell, R. Timothy Hunt, and Paul M. Nurse won the 2001 Nobel Prize in Physiology or Medicine for their discovery of these critical proteins.
What makes a Cell Cancerous?
Cancer is a disease characterized by a population of cells that grow and divide without respect to normal limits. These cancerous cells invade and destroy adjacent tissues, and they may spread throughout the body. The process by which normal cells are transformed into cancer cells is known as carcinogenesis. This process is also known as oncogenesis or tumorigenesis.
Nearly all cancers are caused by mutations in the DNA of abnormal cells. These mutations may be due to the effects of carcinogens, cancer-causing agents such as tobacco smoke, radiation, chemicals, or infectious agents. These carcinogens may act as an environmental “trigger,” stimulating the onset of cancer in certain individuals and not others. Do all people who smoke get cancer? No. Can secondhand smoke increase a nonsmoking person's chance of developing lung cancer? Yes. It also increases a nonsmoking person's chance of developing heart disease.
Complex interactions between carcinogens and an individual’s genome may explain why only some people develop cancer after exposure to an environmental trigger and others do not. Do all cancers need an environmental trigger to develop? No. Cancer-causing mutations may also result from errors incorporated into the DNA during replication, or they may be inherited. Inherited mutations are present in all cells of the organism.
Oncogenes and Tumor Suppressor Genes
Some types of cancer occur because of mutations in genes that control the cell cycle. Cancer-causing mutations most often occur in two types of regulatory genes, called proto-oncogenes and tumor-suppressor genes.
- Proto-oncogenes are genes that normally help cells divide. When a proto-oncogene mutates to become an oncogene, it is continuously active, even when it is not supposed to be. This is like a car's accelerator pedal being stuck at full throttle. The car keeps racing at top speed. In the case of a cell, the cell keeps dividing out of control, which can lead to cancer.
- Tumor suppressor genes are genes that normally slow down or stop cell division. When a mutation occurs in a tumor suppressor gene, it can no longer control cell division. This is like a car without brakes. The car can't be slowed or stopped. In the case of a cell, the cell keeps dividing out of control, which can lead to cancer.
Several Mutations to Cause Cancer
Oncogenes may be growth factors, protein kinases, GTPases or transcription factors. Growth factors are naturally occurring substances, usually a protein or steroid hormone, capable of stimulating cellular growth, proliferation, and differentiation. They are important for regulating a variety of cellular processes. Usually, they must bind to an extracellular or intracellular receptor to initiate a cellular reaction.
Typically, a series of several mutations that constitutively activate oncogenes and inactivate tumor suppressor genes is required to transform a normal cell into a cancer cell (Figure \(\PageIndex{2}\)). Cells have developed a number of control mechanisms to overcome mutations in proto-oncogenes. Therefore, a cell needs multiple mutations to transform into a cancerous cell. A mutation in one proto-oncogene would not cause cancer, as the effects of the mutation would be masked by the normal control of the cell cycle and the actions of tumor suppressor genes. Similarly, a mutation in one tumor suppressor gene would not cause cancer either, due to the presence of many "backup" genes that duplicate its functions. It is only when enough proto-oncogenes have mutated into oncogenes and enough tumor suppressor genes have been deactivated that the cancerous transformation can begin. Signals for cell growth overwhelm the signals for growth regulation, and the cell quickly spirals out of control. Often, because many of these genes regulate the processes that prevent most damage to the genes themselves, DNA damage accumulates as one ages.
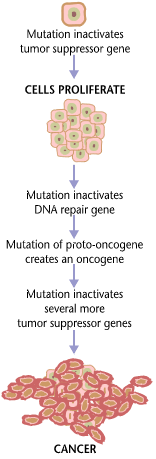
Usually, oncogenes are dominant alleles, as they contain gain-of-function mutations. The actions of the mutant allele gene product, many times resulting in a constitutively activated protein, are dominant to the gene product produced by the "normal" allele. Meanwhile, mutated tumor suppressors are generally recessive alleles, as they contain loss-of-function mutations. A proto-oncogene needs only a mutation in one copy of the gene to generate an oncogene; a tumor suppressor gene needs a mutation in both copies of the gene to render both products defective. There are instances when, however, one mutated allele of a tumor suppressor gene can render the other copy non-functional. These instances result in what is known as a dominant negative effect.
Review
- Define cancer.
- What are cyclin-dependent kinases? What is their role?
- Discuss the role of oncogenes and tumor suppressor genes in carcinogenesis.
- Why are multiple mutations required for transformation into a cancerous cell?
- Identify all the categories of oncogenes and describe two categories.
Attributions
- Cell cycle by WassermanLab, CC BY-SA 4.0, via Wikimedia Commons
- Cancer requires multiple mutations by National Cancer Institute, public domain via Wikimedia Commons
- Text adapted from Human Biology by CK-12 licensed CC BY-NC 3.0