
32.8: Pollination and Fertilization - Development of the Seed
- Page ID
- 13808

- Name the three parts of a seed and describe their functions and development
Parts of a Seed
The seed, along with the ovule, is protected by a seed coat that is formed from the integuments of the ovule sac. In dicots, the seed coat is further divided into an outer coat, known as the testa, and inner coat, known as the tegmen. The embryonic axis consists of three parts: the plumule, the radicle, and the hypocotyl. The portion of the embryo between the cotyledon attachment point and the radicle is known as the hypocotyl. The embryonic axis terminates in a radicle, which is the region from which the root will develop.
Seed Growth
In angiosperms, the process of seed development begins with double fertilization and involves the fusion of the egg and sperm nuclei into a zygote. The second part of this process is the fusion of the polar nuclei with a second sperm cell nucleus, thus forming a primary endosperm. Right after fertilization, the zygote is mostly inactive, but the primary endosperm divides rapidly to form the endosperm tissue. This tissue becomes the food the young plant will consume until the roots have developed after germination. The seed coat forms from the two integuments or outer layers of cells of the ovule, which derive from tissue from the mother plant: the inner integument forms the tegmen and the outer forms the testa. When the seed coat forms from only one layer, it is also called the testa, though not all such testae are homologous from one species to the next.
In gymnosperms, the two sperm cells transferred from the pollen do not develop seed by double fertilization, but one sperm nucleus unites with the egg nucleus and the other sperm is not used. Sometimes each sperm fertilizes an egg cell and one zygote is then aborted or absorbed during early development. The seed is composed of the embryo and tissue from the mother plant, which also form a cone around the seed in coniferous plants such as pine and spruce. The ovules after fertilization develop into the seeds.
Food Storage in the Seed
The storage of food reserves in angiosperm seeds differs between monocots and dicots. In monocots, the single cotyledon is called a scutellum; it is connected directly to the embryo via vascular tissue. Food reserves are stored in the large endosperm. Upon germination, enzymes are secreted by the aleurone, a single layer of cells just inside the seed coat that surrounds the endosperm and embryo. The enzymes degrade the stored carbohydrates, proteins, and lipids. These products are absorbed by the scutellum and transported via a vasculature strand to the developing embryo.
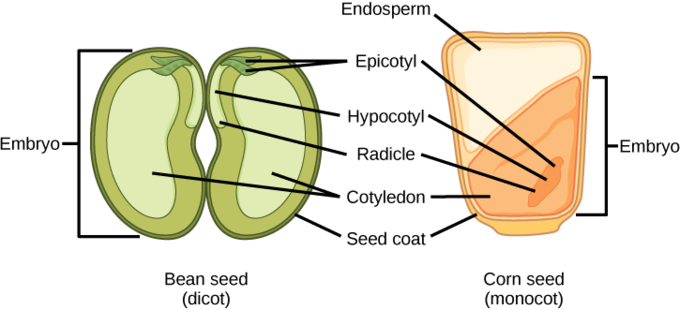
In endospermic dicots, the food reserves are stored in the endosperm. During germination, the two cotyledons act as absorptive organs to take up the enzymatically-released food reserves, similar to the process in monocots. In non-endospermic dicots, the triploid endosperm develops normally following double fertilization, but the endosperm food reserves are quickly remobilized, moving into the developing cotyledon for storage.
Seed Germination
Upon germination in dicot seeds, the epicotyl is shaped like a hook with the plumule pointing downwards; this plumule hook persists as long as germination proceeds in the dark. Therefore, as the epicotyl pushes through the tough and abrasive soil, the plumule is protected from damage. Upon exposure to light, the hypocotyl hook straightens out, the young foliage leaves face the sun and expand, and the epicotyl continues to elongate. During this time, the radicle is also growing and producing the primary root. As it grows downward to form the tap root, lateral roots branch off to all sides, producing the typical dicot tap root system.

In monocot seeds, the testa and tegmen of the seed coat are fused. As the seed germinates, the primary root emerges, protected by the root-tip covering: the coleorhiza. Next, the primary shoot emerges, protected by the coleoptile: the covering of the shoot tip. Upon exposure to light, elongation of the coleoptile ceases and the leaves expand and unfold. At the other end of the embryonic axis, the primary root soon dies, while other, adventitious roots emerge from the base of the stem. This produces the fibrous root system of the monocot.
Depending on seed size, the time it takes a seedling to emerge may vary. However, many mature seeds enter a period of dormancy marked by inactivity or extremely-low metabolic activity. This period may last for months, years, or even centuries. Dormancy helps keep seeds viable during unfavorable conditions. Upon a return to optimal conditions, seed germination takes place. These conditions may be as diverse as moisture, light, cold, fire, or chemical treatments. Scarification, the softening of the seed coat, presoaking in hot water, or passing through an acid environment, such as an animal’s digestive tract, may also be needed.
Key Points
- In angiosperms, the process of seed production begins with double fertilization while in gymnosperms it does not.
- In both monocots and dicots, food reserves are stored in the endosperm; however, in non-endospermic dicots, the cotyledons act as the storage.
- In a seed, the embryo consists of three main parts: the plumule, the radicle, and the hypocotyl.
- In dicots, the hypocotyls extend above ground, giving rise to the stem of the plant, while in monocots, they remain below ground.
- In dicot seeds, the radicle grows downwards to form the tap root while lateral roots branch off to all sides, producing a dicot tap root system; in contrast, the end of germination in monocot seeds is marked by the production of a fibrous root system where adventitious roots emerge from the stem.
- Seed germination is dependent on seed size and whether or not favorable conditions are present.
Key Terms
- testa: the seed coat
- radicle: the rudimentary shoot of a plant that supports the cotyledons in the seed and from which the root is developed downward; the root of the embryo
- hypocotyl: in plants with seeds, the portion of the embryo or seedling between the root and cotyledons
- plumule: consisting of the apical meristem and the first true leaves of the young plant
- coleoptile: a pointed sheath that protects the emerging shoot in monocotyledons such as oats and grasses