23.3B: Chromalveolata- Alveolates
- Page ID
- 13587

Alveolates are defined by the presence of an alveolus beneath the cell membrane and include dinoflagellates, apicomplexans and ciliates.
- Evaluate traits associated with protists classified as alveolates which include dinoflagellates, apicomplexans, and ciliates
Key Points
- Alveolates are classified under the group Chromalveolata which developed as a result of a secondary endosymbiotic event.
- Dinoflagellates are defined by their flagella structure which lays perpendicular and fits into the cellulose plates of the dinoflagellate, promoting a spinning motion.
- Apicomplexans are defined by the asymmetrical distribution of their microtubules, fibrin, and vacuoles; they include the parasitic protist Plasmodium which causes malaria.
- Ciliates are defined by the presence of cilia (such as the oral groove in the Paramecium), which beat synchronously to aid the organism in locomotion and obtaining nutrients.
- Ciliates are defined by the presence of cilia, which beat synchronously, to aid the organism in locomotion and obtaining nutrients, such as the oral groove in the Paramecium.
Key Terms
- osmoregulation: the homeostatic regulation of osmotic pressure in the body in order to maintain a constant water content
- plastid: any of various organelles found in the cells of plants and algae, often concerned with photosynthesis
- conjugation: the temporary fusion of organisms, especially as part of sexual reproduction
Chromalveolata
Current evidence suggests that species classified as chromalveolates are derived from a common ancestor that engulfed a photosynthetic red algal cell, which itself had already evolved chloroplasts from an endosymbiotic relationship with a photosynthetic prokaryote. Therefore, the ancestor of chromalveolates is believed to have resulted from a secondary endosymbiotic event. However, some chromalveolates appear to have lost red alga-derived plastid organelles or lack plastid genes altogether. Therefore, this supergroup should be considered a hypothesis-based working group that is subject to change and can be subdivided into alveolates and stramenopiles.
Alveolates
A large body of data supports that the alveolates are derived from a shared common ancestor. The alveolates are named for the presence of an alveolus, or membrane-enclosed sac, beneath the cell membrane. The exact function of the alveolus is unknown, but it may be involved in osmoregulation. The alveolates are further categorized into the dinoflagellates, the apicomplexans, and the ciliates.
Dinoflagellates
Dinoflagellates exhibit extensive morphological diversity and can be photosynthetic, heterotrophic, or mixotrophic. Many dinoflagellates are encased in interlocking plates of cellulose with two perpendicular flagella that fit into the grooves between the cellulose plates. One flagellum extends longitudinally and a second encircles the dinoflagellate. Together, the flagella contribute to the characteristic spinning motion of dinoflagellates. These protists exist in freshwater and marine habitats; they are a component of plankton.
Some dinoflagellates generate light, called bioluminescence, when they are jarred or stressed. Large numbers of marine dinoflagellates (billions or trillions of cells per wave) can emit light and cause an entire breaking wave to twinkle or take on a brilliant blue color. For approximately 20 species of marine dinoflagellates, population explosions (called blooms) during the summer months can tint the ocean with a muddy red color. This phenomenon is called a red tide and results from the abundant red pigments present in dinoflagellate plastids. In large quantities, these dinoflagellate species secrete an asphyxiating toxin that can kill fish, birds, and marine mammals. Red tides can be massively detrimental to commercial fisheries; humans who consume these protists may become poisoned.

Apicomplexans
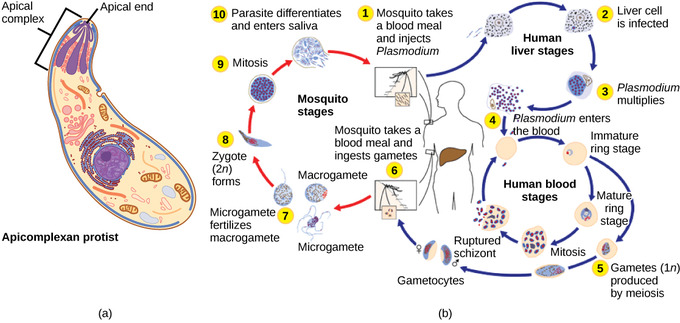
The apicomplexan protists are so named because their microtubules, fibrin, and vacuoles are asymmetrically distributed at one end of the cell in a structure called an apical complex. The apical complex is specialized for entry and infection of host cells. Indeed, all apicomplexans are parasitic. This group includes the genus Plasmodium, which causes malaria in humans. Apicomplexan life cycles are complex, involving multiple hosts and stages of sexual and asexual reproduction.
Ciliates
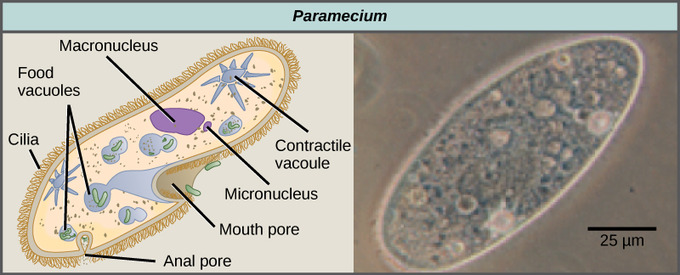
The ciliates, which include Paramecium and Tetrahymena, are a group of protists 10 to 3,000 micrometers in length that are covered in rows, tufts, or spirals of tiny cilia. By beating their cilia synchronously or in waves, ciliates can coordinate directed movements and ingest food particles. Certain ciliates have fused cilia-based structures that function like paddles, funnels, or fins. Ciliates also are surrounded by a pellicle, providing protection without compromising agility. The genus Paramecium includes protists that have organized their cilia into a plate-like primitive mouth called an oral groove, which is used to capture and digest bacteria. Food captured in the oral groove enters a food vacuole where it combines with digestive enzymes. Waste particles are expelled by an exocytic vesicle that fuses at a specific region on the cell membrane: the anal pore. In addition to a vacuole-based digestive system, Paramecium also uses contractile vacuoles: osmoregulatory vesicles that fill with water as it enters the cell by osmosis and then contract to squeeze water from the cell.
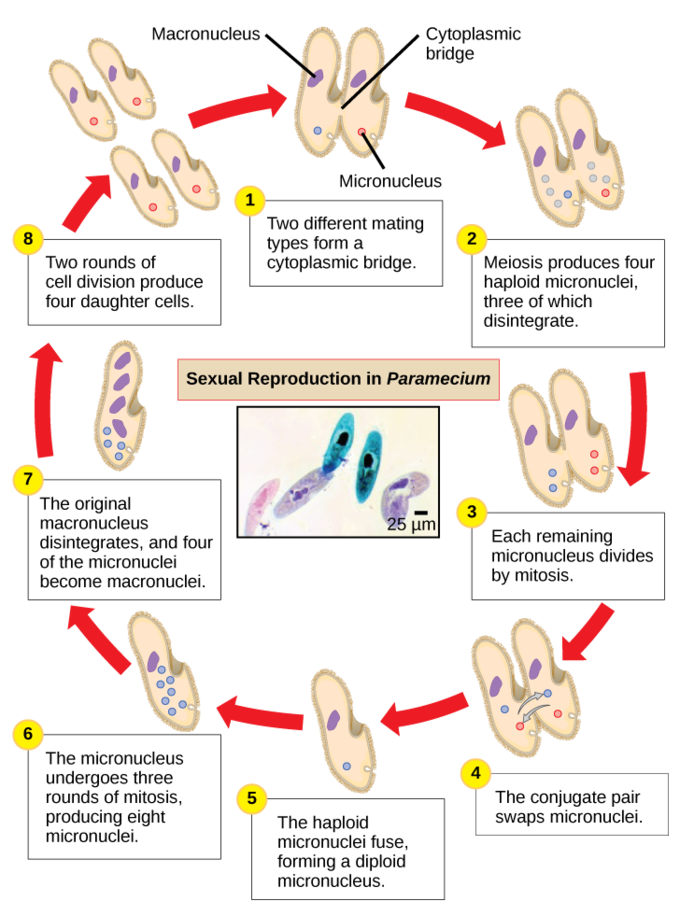
Paramecium has two nuclei, a macronucleus and a micronucleus, in each cell. The micronucleus is essential for sexual reproduction, whereas the macronucleus directs asexual binary fission and all other biological functions. The process of sexual reproduction in Paramecium underscores the importance of the micronucleus to these protists. Paramecium and most other ciliates reproduce sexually by conjugation. This process begins when two different mating types of Paramecium make physical contact and join with a cytoplasmic bridge. The diploid micronucleus in each cell then undergoes meiosis to produce four haploid micronuclei. Three of these degenerate in each cell, leaving one micronucleus that then undergoes mitosis, generating two haploid micronuclei. The cells each exchange one of these haploid nuclei and move away from each other. A similar process occurs in bacteria that have plasmids. Fusion of the haploid micronuclei generates a completely novel diploid pre-micronucleus in each conjugative cell. This pre-micronucleus undergoes three rounds of mitosis to produce eight copies, while the original macronucleus disintegrates. Four of the eight pre-micronuclei become full-fledged micronuclei, whereas the other four perform multiple rounds of DNA replication and then become new macronuclei. Two cell divisions then yield four new paramecia from each original conjugative cell.