
16.15: Regulating Gene Expression in Cell Development - Establishing Body Axes during Development
- Page ID
- 13371

- Summarize the mechanisms and cell types that establish the body axes
Vertebrate axis formation
During development, as the germ layers form, the ball of cells still retains its spherical shape. However, animal bodies have lateral-medial (left-right), dorsal-ventral (back-belly), and anterior-posterior (head-feet) axes. How are these established? In one of the most seminal experiments ever to be carried out in developmental biology, Spemann and Mangold took dorsal cells from one embryo and transplanted them into the belly region of another embryo. They found that the transplanted embryo now had two notochords: one at the dorsal site from the original cells and another at the transplanted site. This suggested that the dorsal cells were genetically programmed to form the notochord and define the axis. Since then, researchers have identified many genes that are responsible for axis formation. Mutations in these genes leads to the loss of symmetry required for organism development. Animal bodies have externally visible symmetry. However, the internal organs are not symmetric. For example, the heart is on the left side and the liver on the right. The formation of the central left-right axis is an important process during development. This internal asymmetry is established very early during development and involves many genes. Research is still ongoing to fully understand the developmental implications of these genes.
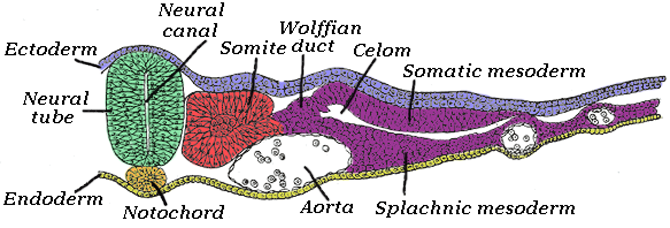
Neural tube
In the developing chordate (including vertebrates), the neural tube is the embryo’s precursor to the central nervous system, which comprises the brain and spinal cord. The neural groove gradually deepens as the neural folds become elevated, and ultimately the folds meet and coalesce in the middle line and convert the groove into a closed tube, the neural tube or neural canal, the ectodermal wall of which forms the rudiment of the nervous system.
Primary and secondary neurulation
The neural tube develops in two ways: primary neurulation and secondary neurulation. Primary neurulation divides the ectoderm into three cell types: the internally located neural tube, the externally located epidermis, and the neural crest cells, which develop in the region between the neural tube and epidermis but then migrate to new locations. Primary neurulation begins after the neural plate forms. The edges of the neural plate start to thicken and lift upward, forming the neural folds. The center of the neural plate remains grounded, allowing a U-shaped neural groove to form. This neural groove sets the boundary between the right and left sides of the embryo. The neural folds pinch in towards the midline of the embryo and fuse together to form the neural tube. In secondary neurulation, the cells of the neural plate form a cord-like structure that migrates inside the embryo and hollows to form the tube.
Each organism uses primary and secondary neurulation to varying degrees. Neurulation in fish proceeds only via the secondary form. In avian species the posterior regions of the tube develop using secondary neurulation and the anterior regions develop by primary neurulation. In mammals, secondary neurulation begins around the 35th somite. Mammalian neural tubes close in the head in the opposite order that they close in the trunk. In the head, neural crest cells migrate, the neural tube closes, and the overlying ectoderm closes. In the trunk, overlying ectoderm closes, the neural tube closes and neural crest cells migrate.
Neural tube subdivisions
Four neural tube subdivisions eventually develop into distinct regions of the central nervous system by the division of neuroepithelial cells: the prosencephalon, the mesencephalon, the rhombencephalon and the spinal cord. The prosencephalon further goes on to develop into the telencephalon (the forebrain or cerebrum) and the diencephalon (the optic vesicles and hypothalamus). The mesencephalon develops into the midbrain. The rhombencephalon develops into the metencephalon (the pons and cerebellum) and the myelencephalon (the medulla oblongata).
For a short time, the neural tube is open both cranially and caudally. These openings, called neuropores, close during the fourth week in the human. Improper closure of the neuropores can result in neural tube defects such as anencephaly or spina bifida. The dorsal part of the neural tube contains the alar plate, which is primarily associated with sensation. The ventral part of the neural tube contains the basal plate, which is primarily associated with motor (i.e., muscle) control.
Signaling molecules and other factors
The neural tube patterns along the dorsal-ventral axis establish defined compartments of neural progenitor cells that lead to distinct classes of neurons. This patterning occurs early in development and results from the activity of several secreted signaling molecules. Sonic hedgehog (Shh) is a key player in patterning the ventral axis, while Bone morphogenic proteins (Bmp) and Wnt family members play an important role in patterning the dorsal axis. Other factors shown to provide positional information to the neural progenitor cells include Fibroblast growth factors (FGF) and Retinoic Acid. Retinoic acid is required ventrally along with Shh to induce Pax6 and Olig2 during differentiation of motor neurons. Three main ventral cell types are established during early neural tube development: the floor plate cells, which form at the ventral midline during the neural fold stage; as well as the more dorsally located motor neurons and interneurons. These cell types are specified by the secretion of Shh from the notochord (located ventrally to the neural tube), and later from the floor plate cells. Shh acts as a morphogen, meaning that it acts in a concentration-dependent manner to specify cell types as it moves further from its source. The different combinations of expression of transcription factors along the dorsal-ventral axis of the neural tube are responsible for creating the identity of the neuronal progenitor cells.
Key Points
- The three axes of the animal body are established in development via the expression of specific sets of genes that regulate which cells will develop into specific structures.
- During development, the dorsal cells are genetically programmed to develop into the notochord and define the axis.
- The neural tube can develop in two ways: primary or secondary neurulation, which are used by organisms in varying degrees to establish the neural tube that will develop into the central nervous system (brain and spinal cord).
- Specific patterns along the neural tube that are established via secretion and production of specific signaling molecules (such as Wnt, Shh, BMP and retinoic acid) play a key role in patterning the dorsal and ventral axes.
Key Terms
- neural tube: hollow longitudinal dorsal tube formed in the folding and subsequent fusion of the opposite ectodermal folds in the embryo that gives rise to the brain and spinal cord
- neurulation: the process by which the beginnings of the vertebrate nervous system is formed in embryos
- anencephaly: a lethal birth defect in which most of the brain and parts of the skull are missing; absence of the encephalon
- notochord: a flexible rodlike structure that forms the main support of the body in the lowest chordates; a primitive spine