
16.4D: Flowering
- Page ID
- 5805

The flowering plants (angiosperms) go through a phase of vegetative growth producing more stems and leaves and a flowering phase where they produce the organs for sexual reproduction. In "annuals", like the snapdragon, the vegetative phase begins with germination of the seed. Flowering follows and ends with the senescence and death of the plant. In biennials, the vegetative phase takes up the first year; flowering followed by death occurs the second year. In perennials, flowering typically occurs year after year when conditions are appropriate.
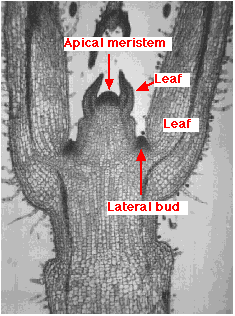
Vegetative growth of the above-ground part of the plant — the shoot — occurs at the apical meristem. This is a mass of undifferentiated cells at the tip of the stem. Mitosis of these cells produces cells that differentiate to form more stem, leaves and secondary meristems. Also called lateral buds, these form in the axils of the leaves and will form branches.
The Signal to Flower
Flowering involves the conversion of the apical meristem into a floral meristem, from which all the parts of the flower will be produced. Signals that change the fate of the apical meristem include:
- maturity of the plant
- temperature
- the arrival of the plant hormone gibberellin
- for many plants, photoperiod - the relative length of day and night.
Temperature
Many annual plants (e.g., winter wheat) and biennial plants have their time of flowering delayed unless they have undergone a preceding period of wintertime cold. The change brought about by this prolonged exposure to the cold is called vernalization.
In the "model" plant Arabidopsis thaliana, vernalization works like this.
- A gene designated Flowering Locus C (FLC) encodes a transcription factor that blocks the expression of the genes needed for flowering.
- The level of FLC mRNA is high in the fall.
- But with the onset of cold temperatures, production of an antisense transcript of FLC (called COOLAIR) increasesas does, later, a sense transcript of part of the FLC gene.
- Both of these RNAs are non-coding; that is, they are not translated into protein.
- But they cooperate in suppressing the production of FLC mRNA and its translation into FLC protein.
- With the arrival of spring, there is no FLC protein remaining to suppress flowering so flowering can begin.

The buds of many species of woody angiosperms found in temperate climates, such as apples and lilacs, also need a preceding period of cold weather before they can develop into flowers. So these plants cannot be grown successfully at lower latitudes because the winters never get cold enough (a few days at 0–10°C). This bud dormancy is localized. Prior chilling of one bud on a lilac stem enables it to flower while the other, nonchilled, buds on the stem remain dormant.
Photoperiod
Photoperiod is detected in the leaves. The cocklebur, drawn here, needs at least 8.5 hours of darkness in order to flower. Even if only a part of one leaf is exposed to the correct photoperiod, the entire plant will bloom (middle figure).
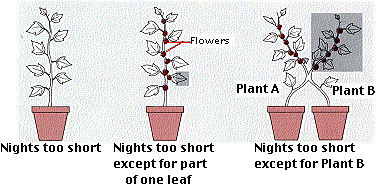
The leaves produce a chemical signal called florigen that is transmitted to the apical meristems to start their conversion into floral meristems. The right-hand drawing shows that grafting a cocklebur (B) that receives the required period of darkness to one (A) that does not causes flowering in both. Evidently the florigen signal passes from B to A through their connected vascular systems.
The chemical nature of florigen has been sought for decades. The most recent evidence suggests that at least one component is the protein encoded by the gene FLOWERING LOCUS T (FT).
Converting the Apical Meristem to a Floral Meristem
In the nucleus of the meristem cells, the FT protein binds to the transcription factor FD and turns on the expression of genes needed for flowering, e.g., APETALA1 and LEAFY.
Structure of the Flower
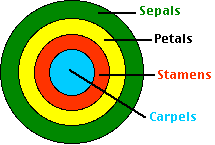
The floral meristem differentiates into four concentric groups of cells that form the four parts of the flower.
- The cells in whorl 1 develop into a whorl of sepals. These form at the lowest level. Collectively they make up the calyx.
- Whorl 2 forms above the calyx, forming the petals. Collectively these make up the corolla of the flower (the part that most ornamentals are grown for).
- Whorl 3 develops into the stamens, the male reproductive organs.
- The innermost whorl, 4, forms carpels, the female reproductive organs. Carpels often fuse to form a single structure, which some botanists call the pistil.
What triggers the various parts of the floral meristem to enter one or another of these four developmental pathways?
The ABC Model of Flower Development
Genetic analysis of mutants especially those found in the dicots Arabidopsis thaliana and in the snapdragon (Antirrhinum) support the ABC model of flowering. This model postulates a group of genes that encode the transcription factors needed to turn on the genes for sepal, petal, etc. development. The "master switches" fall into 3 groups: A, B, and C.
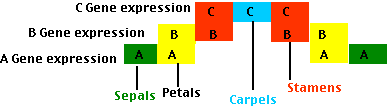
These are the rules:
- Cells in which only A genes are expressed develop into sepals. This occurs at the lowest level of the floral meristem.
- Cells in which both A and B genes are expressed develop into petals. This occurs at the next higher level.
- Expression of B and C genes turns on the developmental program to form stamens.
- Expression of C genes alone turns on the development of carpels in the innermost band of cells.
Examples of A, B and C group genes involved in flowering - these have been identified in Arabidopsis thaliana
| A group | APETALA1 (AP1) and APETALA2 (AP2) |
| B group | APETALA3 (AP3) and PISTILLATA (PI) |
| C group | AGAMOUS (AG) |
The transcription factor LEAFY plays a major role in turning on the A, B, and C group genes in the appropriate locations.
- The LEAFY protein alone turns on AP1 in whorls 1 and 2.
- LEAFY plus a protein encoded by the gene UFO (for "unusual floral organs") turn on AP3 in whorls 2 and 3.
- LEAFY and a second, still unidentified, protein turn on AG in whorls 3 and 4.
If LEAFY protein alone is sufficient to turn on AP1, why isn't AP1 expressed in all four whorls?
The answer: AGAMOUS blocks the expression of AP1, so any cell expressing AGAMOUS cannot express AP1. In fact, the antagonism is reciprocal: AP2 in whorls 1 and 2 (A group) inhibits AGAMOUS so the gene expression in whorls 3 and 4 remains distinct from that in whorls 1 and 2.
The proteins encoded by APETALA3 and PISTILLATA (Group B) form a heterodimer that binds to
- the APETALA1 protein to form petals
- the AGAMOUS protein to form stamens
Aided by a fourth transcription factor encoded by the gene SEPALLATA3, these quaternary complexes bind to specific sequences of DNA turning on the expression of the various genes needed to form whorls 2 and 3. Further research may reveal similar behavior for the other genes.
SEPALLATA3 (SEP3) is one of four SEP genes in Arabidopsis. If all but SEP4 are inactivated, a flower with only sepals is formed (hence the name). If all four are inactivated, no flowers are formed at all.
So formation of a flower requires a cascade of sequential gene activity that gradually converts a mass of undifferentiated cells (the apical meristem) into the parts of a flower. The genes encode transcription factors that act as master switches, turning on (or off) downstream genes that ultimately make each part of the flower in its appropriate location. This same strategy of genetic control of developmental pathways is seen in animal development.