
15.4M: Antigen Presentation
- Page ID
- 5449

Antigens are macromolecules that elicit an immune response in the body. Antigens can be proteins, polysaccharides, conjugates of lipids with proteins (lipoproteins) and polysaccharides (glycolipids). Most of this page will describe how protein antigens are presented to the immune system. The presentation of lipid and polysaccharide antigens will be mentioned at the end. It will be helpful to distinguish between two limiting cases.
Antigens that enter the body from the environment; these would include inhaled macromolecules (e.g., proteins on cat hairs that can trigger an attack of asthma in susceptible people), ingested macromolecules (e.g., shellfish proteins that trigger an allergic response in susceptible people), and molecules that are introduced beneath the skin (e.g., on a splinter or in an injected vaccine). Alternatively, antigens can be generated within the cells of the body; these would include proteins encoded by the genes of viruses that have infected a cell and aberrant proteins that are encoded by mutant genes; such as mutated genes in cancer cells. In all cases, however, the initial immune response to any antigen absolutely requires that the antigen be recognized by a T lymphocyte ("T cell"). The truth of this rule is clearly demonstrated in AIDS: the infections (viral or fungal or bacterial) that so often claim the life of AIDS patients do so when the patient has lost virtually all of his or her CD4+ T cells. The two categories of antigens are processed and presented to T cells by quite different mechanisms.
Exogenous Antigens
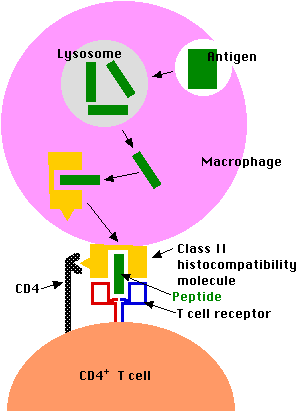
Exogenous antigens (inhaled, ingested, or injected) are taken up by antigen-presenting cells (APCs). These include phagocytic cells like dendritic cells and macrophages and B lymphocytes ("B cells") which are responsible for producing antibodies against the antigen. Antigen-presenting cells
- engulf the antigen by endocytosis
- endosome fuses with a lysosome where the antigen is degraded into fragments (e.g. short peptides)
- these antigenic peptides are then displayed at the surface of the cell nestled within a class II histocompatibility molecule.
- they may be recognized by CD4+T cells
The Class I Pathway
Class I histocompatibility molecules are transmembrane proteins expressed at the cell surface. Like all transmembrane proteins, they are synthesized by ribosomes on the rough endoplasmic reticulum (RER) and assembled within its lumen. There are three subunits in each class I histocompatibility molecule:
- the transmembrane polypeptide (called the "heavy chain")
- the antigenic peptide
- beta-2 microglobulin
All of these must be present within the lumen of the endoplasmic reticulum if they are to assemble correctly and move through the Golgi apparatus to the cell surface. The Problem: proteins encoded by the genes of an infecting virus are synthesized in the cytosol. How to get them into the endoplasmic reticulum?
The Solution: TAP (= transporter associated with antigen processing).
- Viral proteins in the cytosol are degraded by proteasomes into viral peptides.
- The peptides are picked up by TAP proteins embedded in the membrane of the endoplasmic reticulum.
- Using the energy of ATP, the peptides are pumped into the lumen of the endoplasmic reticulum where they assemble with the transmembrane polypeptide and beta-2 microglobulin.
- This trimolecular complex then moves through the Golgi apparatus and is inserted in the plasma membrane.
- The complex can be bound by a T cell with a receptor (TCR) able to bind the peptide and flanking portions of the histocompatibility molecule (the hot dog in the bun) and CD8 molecules that bind the CD8 receptor (shown above as a gray hemisphere) on the histocompatibility molecule.
The Class II Pathway
Class II histocompatibility molecules consist of two transmembrane polypeptides and a third molecule nestled in the groove they form. All three components of this complex must be present in the endoplasmic reticulum for proper assembly. But antigenic peptides are not transported to the endoplasmic reticulum, so a protein called the invariant chain ("Ii") temporarily occupies the groove.
The steps:
- The two chains of the class II molecule are inserted into the membrane of the endoplasmic reticulum.
- They bind (in their groove) one molecule of invariant chain.
- This trimolecular complex is transported through the Golgi apparatus and into vesicles called lysosomes.
Meanwhile foreign antigenic material is engulfed by endocytosis forming endosomes. These also fuse with lysosomes. Then,
- The antigen is digested into fragments.
- The invariant (Ii) chain is digested.
- This frees the groove for occupancy by the antigenic fragment.
- The vesicles move to the plasma membrane and the complex is displayed at the cell surface.
- The complex can be bound by a T cell with
- a receptor (TCR) able to bind the peptide and flanking portions of the histocompatibility molecule (the hot dog in the bun) and
- CD4 molecules that bind the CD4 receptor (shown above as a yellow triangle) found on all class II histocompatibility molecules.
Interconnections Between the Class I and Class II Pathways
Cross-Presentation: Transferring Exogenous Antigens to the Class I Pathway
Cross-presentation is the transferring of extracellular antigens like bacteria, some tumor antigens, and antigens in cells infected by viruses into the class I pathway for stimulation of CD8+ cytotoxic T cells (CTL). Only certain "professional" antigen-presenting cells (APCs) like dendritic cells can do this - use the class I as well as the class II pathways of antigen presentation.
Cross-presentation following infection by viruses is important because:
- Most viruses infect cells other than APCs (e.g., liver cells, epithelial cells of the lung) (and, of course, are intracellular in these).
- While viral antigens displayed on the surface of any infected cell can serve as targets for cytotoxic T cells (CTLs),
- the lack of any costimulatory molecules on the cell surface makes them poor stimulants for the development of clones of CTLs in the first place.
However, when an infected cell dies, it can be engulfed by a professional APC, and the antigens within it can enter the class I pathway. One mechanism:
- The dead cell is engulfed by endocytosis.
- The endosome that forms fuses with a lysosome and degradation of the dead cell begins.
- Antigens pass into the cytosol and are degraded in proteasomes.
- The peptides formed are then are picked up by TAP and inserted into class I MHC molecules and displayed at the cell surface — along with the costimulatory molecules needed to start a vigorous clonal expansion of CD8+ cytotoxic T cells.
Diverting Antigens from the Class I to the Class II Pathway
Autophagy provides a mechanism by which cells can transfer endogenous (intracellular) antigens into the class II pathway, for example
- self-proteins so as to be able to delete CD4+ T cells with receptors capable of attacking them and thus potentially capable of causing autoimmunity
- proteins synthesized by an infecting virus. In this way viral infection can generate CD4+ T cells as well as cytotoxic T cells (CD8+)
B Lymphocytes: A Special Case
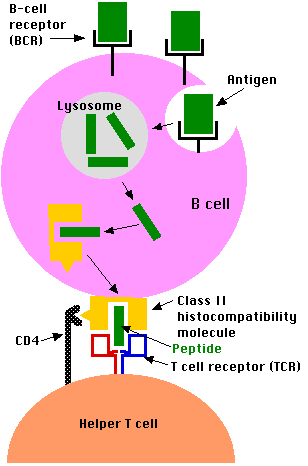
B lymphocytes are both antigen-receiving and antigen-presenting cells. They bind intact antigens (e.g., virus particles, proteins) with their B cell receptor (BCR). They can come in contact with these antigens by encountering them in the surrounding lymph or by being presented them by macrophages or dendritic cells. B lymphocytes process antigen by the class II pathway for presentation to T cells.
The process:
- B cells engulf antigen by receptor-mediated endocytosis
- The B cell receptors for antigen (BCRs) are antibodies anchored in the plasma membrane.
- The affinity of these for an epitope on an antigen may be so high that the B cell can bind and internalize the antigen when it is present in body fluids in concentrations thousands of times smaller than a macrophage would need.
- The remaining steps of antigen processing occur by the same class II pathway described above for macrophages producing
- fragments of antigen displayed at the cell surface nestled in the groove of class II histocompatibility molecules.
- A CD4+ T cell that recognizes the displayed antigen is stimulated to release lymphokines.
- These, in turn, stimulate the B cell to enter the cell cycle.
- Because of the part they play in stimulating B cells, these CD4+ T cells are called Helper T cells ("Th").
- The B cell grows into a clone of cells (called plasma cells)
- These synthesize receptors (BCRs) with the identical binding site for the epitope but without the transmembrane tail.
- These antibodies are secreted into the surroundings.
Lipid and Polysaccharide Antigens
Lipid Antigens
- Lipid antigens are presented to T cells by cell-surface molecules designated CD1 ("cluster of differentiation" 1).
- Antigen-presenting cells express several different forms of CD1 at their surface. Each is probably specialized to bind a particular type of lipid antigen (e.g. lipopeptide vs glycolipid).
- The exposed surface of CD1 molecules forms an antigen-binding groove much like that of MHC molecules except that
- the amino acids in the groove are more hydrophobic than those in MHC molecules.
- Like protein antigens, lipid antigens are also presented as fragments, i.e., as a "hot dog in a bun".
Polysaccharide Antigens
Some bacterial polysaccharides ingested by APCs
- can be degraded in their lysosomes
- and presented to T cells by MHC class II molecules.
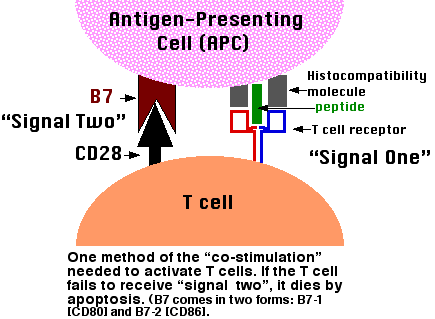
The binding of a T cell to an antigen-presenting cell (APC) is by itself not enough to activate the T cell and turn it into an effector cell: one able to, for examples,
- kill the APC (CD8+ cytotoxic T lymphocytes [CTLs])
- carry out cell-mediated immune reactions (CD4+ Th1 cells)
- provide help to B cells (CD4+ Th2 cells)
In order to become activated, the T cell must not only bind to the epitope (MHC-peptide) with its TCR but also receive a second signal from the APC. The receipt of this second signal is called costimulation. Among the most important of these costimulators are molecules on the APC designated B7 and their ligand on the T cell designated CD28. The binding of CD28 to B7 provides the second signal needed to activate the T cell.
Although T cells may encounter self antigens in body tissues, they will not respond unless they receive a second signal. In fact, binding of their TCR ("signal one") without "signal two" causes them to self-destruct by apoptosis. Most of the time, the cells presenting the body's own antigens either
- fail to provide signal two or
- transmit an as-yet-unidentified second signal that turns the T cell into a regulatory T cell (Treg) that suppresses immune responses.
In either case, self-tolerance results.