
12.1: Cancer in General
- Page ID
- 5076

A cancer is an uncontrolled proliferation of cells. In some the rate is fast; in others, slow; but in all cancers the cells never stop dividing. This distinguishes cancers — malignant tumors — from benign growths like moles where their cells eventually stop dividing (usually). Even more important, benign growths differ from malignant ones in not producing metastases; that is, they do not seed new growths elsewhere in the body.
Cancers are clones. No matter how many trillions of cells are present in the cancer, they are all descended from a single ancestral cell. Evidence: Although normal tissues of a woman are a mosaic of cells in which one X chromosome or the other has been inactivated, all her tumor cells — even if from multiple sites — have the same X chromosome inactivated.
Cancers begin as a primary tumor. Most (maybe all) solid tumors shed cells into the lymph and blood. Most of these lack the potential to develop into tumors. However, some of the shed cells are able to take up residence and establish secondary tumors — metastases — in other locations of the body. These metastases, not the primary tumor, are what usually kills the patient.
Cancer cells are usually less differentiated than the normal cells of the tissue where they arose. Many people feel that this reflects a process of dedifferentiation, but I doubt it. Rather, evidence is accumulating that cancers arise in precursor cells — stem cells or "progenitor cells" — of the tissue: cells that are dividing by mitosis producing daughter cells that are not yet fully differentiated.
A cancer is an uncontrolled proliferation of cells.
Cancer is a Genetic Disease
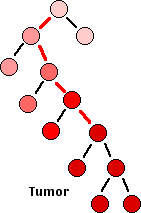
What probably happens is:
- A single cell — perhaps an adult stem cell or progenitor cell — in a tissue suffers a mutation (red line) in a gene involved in the cell cycle, e.g., an oncogene or tumor suppressor gene.
- This results in giving that cell a slight growth advantage over other dividing cells in the tissue.
- As that cell develops into a clone, some if its descendants suffer another mutation (red line) in another cell-cycle gene.
- This further deregulates the cell cycle of that cell and its descendants.
- As the rate of mitosis in that clone increases, the chances of further DNA damage increases.
- Eventually, so many mutations have occurred that the growth of that clone becomes completely unregulated.
- The result: full-blown cancer. (Genetic analysis reveals an average of 63 mutations in pancreatic cancers; almost as many in one type of adult brain cancer, but only 11 somatic mutations in a case of brain cancer in a child.)
- Sequencing samples from several areas in a primary tumor, as well as from some of its metastases, reveals a different collection of mutations from sample to sample. This finding is reinforced by the sequencing of the genome of individual cells from a single tumor each of which shows a unique pattern of shared and unique mutations. (The ability to sequence the genome of a single cell reveals that even normal cells in an adult have accumulated a suite of somatic mutations that differs from cell to cell. However, the rate of somatic mutations in these normal cells is only a fourth of that in cancer cells.)
So even though all the malignant cells in a cancer are descended from a single original cell — and thus are members of a single clone — they are no longer genetically-identical. As the tumor develops, its various cells develop a variety of additional mutations, and these give rise to "subclones" of varying degrees of malignancy with varying
- propensity to metastasize;
- susceptibility to treatment by anticancer drugs;
- propensity to relapse after apparently-successful therapy.
These findings should stimulate a reexamination of the use of chemotherapy.
- While chemotherapy may wipe out dominant subclones in a tumor, there is evidence that is also exerts a selective pressure for the expansion of more malignant, previously-minor, subclones.
- Most chemotherapeutic agents damage DNA so while killing off some cells, they will raise the mutation rate in any surviving cells perhaps encouraging the outgrowth of even more malignant subclones.
Evidence: In a group of patients with chronic lymphocytic leukemia, those receiving chemotherapy survived for shorter periods than those that did not.
Cancer Stem Cells
Stem cells are cells that divide by mitosis to form either two stem cells, thus increasing the size of the stem cell "pool", or one daughter that goes on to differentiate, and one daughter that retains its stem-cell properties. There is growing evidence that most of the cells in leukemias, breast, brain, skin, ovarian, and colon cancers are not able to proliferate out-of-control (and to metastasize). Only those members of the clone that retain their stem-cell-like properties (~2.5% of the cells in a tumor of the colon) can do so.
There is a certain logic to this. Most terminally-differentiated cells have limited potential to divide by mitosis and, seldom passing through S phase of the cell cycle, are limited in their ability to accumulate the new mutations that predispose to becoming cancerous. Furthermore, they often have short life spans — being eliminated by apoptosis (e.g., lymphocytes) or being shed from the tissue (e.g., epithelial cells of the colon). The adult stem cell pool, in contrast, is long-lived, and its members have many opportunities to acquire new mutations as they produce differentiating daughters as well as daughters that maintain the stem cell pool.
Colon cancer
- Begins with the development of polyps in the epithelium of the colon. Polyps are benign growths.
- As time passes, the polyps may get bigger.
- At some point, nests of malignant cells may appear within the polyps
- If the polyp is not removed, some of these malignant cells will escape from the primary tumor and metastasize throughout the body.

Examination of the cells at the earliest, polyp, stage, reveals that they contain one or two mutations associated with cancer. Frequently these include
- the deletion of a healthy copy of the APC (adenomatous polyposis coli) gene on chromosome 5 leaving behind a mutant copy of this tumor suppressor gene
Two results:
- One of the functions of the APC gene product is to destroy the transcription factor β-catenin thus preventing it from turning on genes that cause the cell to divide. With no, or a defective, APC protein, the normal brakes on cell division are lifted.
- Another function of the APC protein is to help attach the microtubules of the mitotic spindle to the kinetochores of the chromosomes. With no, or a defective, APC product available, chromosomes are lost from the spindle producing aneuploid progeny.
- a mutant oncogene (often RAS).
- deletion and/or mutation of the tumor suppressor gene p53
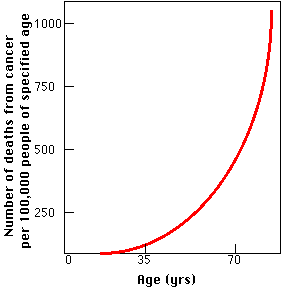
The graph also explains why cancer has become such a common cause of death during the twentieth century. It probably has very little to do with exposure to the chemicals of modern living and everything to do with the increased longevity that has been such a remarkable feature of the 20th century. A population whose members increasingly survive accidents and infectious disease is a population increasingly condemned to death from such "organic" diseases as cancer.
Causes of Cancer
Cancers are caused by
- anything that damages DNA; that is anything that is mutagenic
- radiation that can penetrate to the nucleus and interact with DNA
- chemicals that can penetrate to the nucleus and damage DNA. Chemicals that cause cancer are called carcinogens.
- anything that stimulates the rate of mitosis. This is because a cell is most susceptible to mutations when it is replicating its DNA during the S phase of the cell cycle.
- certain hormones (e.g., hormones that stimulate mitosis in tissues like the breast and the prostate gland)
- chronic tissue injury (which increases mitosis in the stem cells needed to repair the damage)
- agents that cause inflammation (which generates DNA-damaging oxidizing agents in the cell)
- certain other chemicals; some the products of technology
- certain viruses
Viruses and Cancer
Many viruses have been studied that reliably cause cancer when laboratory animals are infected with them. What about humans? The evidence obviously is indirect but some likely culprits are:
- two papilloma viruses that can cause cancer of the cervix and other regions of the genitals (male as well as female).
- the hepatitis B and hepatitis C viruses, which infect the liver and are closely associated with liver cancer (probably because of the chronic inflammation they produce)
- some herpes viruses such as the Epstein-Barr virus (implicated in Burkitt's lymphoma) and KSHV that is associated with Kaposi's sarcoma (a malignancy frequently seen in the late stages of AIDS)
- two human T-cell lymphotropic viruses, HTLV-1 and HTLV-2
But note that the viral infection only contributes to the development of cancer.
- Many people are infected by these viruses and do not develop cancer.
- When cancers do arise in infected people, they still follow our rule of clonality. Many cells have been infected, but only one (usually) develops into a tumor.
So again it appears that only if an infected cell is unlucky enough to suffer several other types of damage will it develop into a tumor.Nevertheless, widespread vaccination against these viruses should not only prevent disease but lower the incidence of the cancers associated with them. A vaccine against hepatitis B is available as are two vaccines (Gardasil® and Cervarix®) against the most dangerous papilloma viruses.
Are Cancers Contagious?
The short answer is NO.
The reason: Cancer cells, like all cells in the body, express histocompatibility molecules on their surface. So like any organ or tissue transplant between two people (other than identical twins), they are allografts and are recognized and destroyed by the recipient's immune system.
However, there are some exceptions.
- Although tumors are not transmissible, viruses are. So any of the viruses described in the previous section can be spread from person to person and predispose them to the relevant cancers.
- There have been a number of cases where, unbeknownst to the surgeon, an organ (e.g., a kidney) from a donor with melanoma has allowed the growth of the same melanoma in the recipient. Transplant recipients must have their immune system suppressed if the transplant is not to be rejected, but their immunosuppression also prevents their immune system from attacking the melanoma cells. Stopping immune suppression cures the recipient (but also causes loss of the kidney).
- Canine transmissible venereal tumor (CTVT). This tumor spreads from dog to dog during copulation. Although the MHC alleles on the tumor cells are only weakly expressed, they do eventually cause the tumor to be rejected.
- Devil facial tumor disease (DFTD). The carnivorous Tasmanian devil is a marsupial living in Tasmania, Australia. The population is threatened by a facial cancer that is spread through bites. The population is highly inbred, thus closely-related genetically, and the MHC alleles on the tumor are only weakly expressed. So it may be these factors that allow the tumor to grow unchecked.
- The soft-shell clam, Mya arenaria, along the North Atlantic coast of North America is being devastated by a leukemia that spreads from animal to animal perhaps as these filter feeders ingest sea water in which leukemic cells have been shed. These mollusks are invertebrates and lack powerful tissue rejection molecules like the MHC of vertebrates.
- There are extremely rare cases where a pregnant woman with cancer (a leukemia or melanoma) has transmitted the cancer across the placenta to her fetus (whose immune system has yet to develop).
The Hallmarks of Cancer
In the year 2000 Douglas Hanahan and Robert Weinberg published a paper — The Hallmarks of Cancer — outlining 6 characteristics that are acquired as a cell progresses toward becoming a full-blown cancer. In the 4 March 2011 issue of Cell, they add 4 other features.
- Uncontrolled proliferation.
- Evasion of growth suppressors. Among the many mutations found in cancers, one or more inactivate tumor suppressor genes.
- Resistance to apoptosis (programmed cell death).
- Develop replicative immortality; i.e., avoid the normal process of cell senescence.
- Induce angiogenesis; that is, promote the development of a blood supply.
- Invasion and metastasis — the ability of tumor cells to invade underlying tissue and then to be carried to other parts of the body where secondary tumors develop (metastasis). During this process, the normal adhesion of cells to each other and to the underlying extracellular matrix (ECM) are disrupted.
- Genomic instability. Cancer cells develop chromosomal aberrations and many (hundreds) of mutations. Most of the latter are "passenger" mutations, but as many as 10 may be "drivers" of the cancerous transformation.
- Inflammation. Tumors are invaded by cells of the immune system, which promote inflammation. One effect of inflammation is the production of reactive oxygen species (ROS). These damage DNA and other molecules.
- Changed energy metabolism. Even if well-supplied with oxygen, cancer cells get most of their ATP from glycolysis not cellular respiration.
- Evade the immune system.