
18.1: Evolution and Adaptation
- Page ID
- 5916

In 1859 the English naturalist Charles Darwin published The Origin of Species. The book contained two major arguments: First, Darwin presented a wealth of evidence of evolution. He said that all living things on earth today are the descendants - with modifications - of earlier species. Second, he proposed a mechanism - natural selection - to explain how evolution takes place.
Evolution involves two interrelated phenomena: adaptation and speciation. In adaptation, over the course of time, species modify their phenotypes in ways that permit them to succeed in their environment. In speciation, over the course of time, the number of species multiplies; that is, a single species can give rise to two or more descendant species. In fact, Darwin maintained that all species are related; that is, any two species on earth today have shared a common ancestor at some point in their history.
Natural Selection
- Living things produce more offspring than the finite resources available to them can support.
- Thus living things face a constant struggle for existence.
- The individuals in a population vary in their phenotypes.
- Some of this variation is inheritable; that is, it is a reflection of variations in genotype.
- Those variants best adapted to the conditions of their life are most likely to survive and reproduce themselves ("survival of the fittest").
- To the extent that their adaptations are inheritable, they will be passed on to their offspring.
The forces of natural selection act on phenotypes, but only if there is a change in the genotypes of a population has evolution occurred.
The Measure of "Fitness"
Fitness is a measure of reproductive success. Those individuals who leave the largest number of mature offspring are the fittest. This can be achieved in several ways:
- Survival (mortality selection)
- Mating success (sexual selection)
- Family size (fecundity selection)
Survival
Any trait that promotes survival - at least until one's reproductive years are over - increases fitness. Such traits are adaptations.
Sexual Selection
In sexual selection, one sex - usually the female - chooses among the available males. Any inherited trait that improves the mating success of certain individuals will become more pronounced in succeeding generations. Some examples:
- When ready to mate, female three-spined sticklebacks (fish) choose males with many Class II MHC alleles over males with fewer alleles. Class II alleles encode the proteins that present antigens to the immune system. Presumably, the more of them you have, the greater the diversity of parasite antigens your immune system can recognize and defend against.The females distinguish between the males by soluble molecules ("odors") the males release into the water. How these "odors" are controlled by the MHC alleles is not known.
- A culture of Drosophila set up with equal numbers of red-eyed and white-eyed flies of both sexes will, after 25 generations or so, end up having only red-eyed (the "normal") flies in it. This despite the fact that white-eyed flies are just as healthy and live just as long as red-eyed flies, i.e., they are equal in terms of survival. But, as it turns out, not only do red-eyed females prefer red-eyed males, but white-eyed females do also.
In other cases of sexual selection, one phenotype prefers to mate with others of the same phenotype. This is called assortative mating.
Fecundity Selection
The production of a large number of mature offspring is a measure of fitness. I stress mature because only they can pass these traits on to another generation. Some ways to do this:
- Earlier breeding. If some females become sexually mature earlier than others, their chances of leaving offspring are enhanced.
- For some species (e.g., fish, oysters), which provide little or no care for their young, fitness is measured by the number of fertilized eggs they produce.
- For species (such as ourselves) that take care of their young, selection acts to reduce family size (to a point). A large study in Utah (U.S.A) showed that in the 19th century, families with fewer children had more surviving grandchildren.
All the forces of natural selection outlined above work on individuals. But there is an increasing body of evidence that natural selection can also act on groups. Natural selection that appears to work counter to the benefit of some individuals while enhancing the prospects of their relatives is called kin selection. It is discussed on a separate page.
Are Humans Exempt from Natural Selection?
It has been argued that advances in medicine, sanitation, etc. have removed humans from the rigors of natural selection. There is probably some truth to this, but consider that of all the human eggs that are fertilized, fewer than half will ever reproduce themselves. The others are eliminated as follows:
- Mortality selection
- Approximately 30% of pregnancies end by spontaneous abortion of embryos and fetuses or by stillbirth.
- Death in infancy and childhood claims another 5% or more.
- Sexual selection
- Another 20% will survive to adulthood but never marry.
- Fecundity selection
- Of those that do marry, 10% will have no children.
Continuous Variation
Most traits in a population such as height and body weight vary in a continuous way from one extreme to the other.

A plot of the distribution of the trait in a population often produces a bell-shaped curve like this one that shows the distribution of heights among a group of male secondary-school seniors. Such a distribution could arise from environmental factors - perhaps the continuous height variation in the boys is simply a result of variation in their diet as they grew up or genetic factors - tall parents tend to have tall children or - most likely - both.
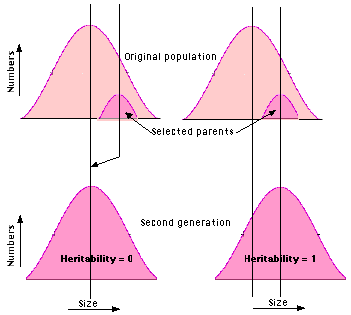
Heritability
One can sort out the relative contribution of genetic and environmental factors by comparing the range of a trait in the offspring compared with the average value of that trait in their parents. If the offspring of selected parents occupy the same range as the entire population, environmental factors are working alone. The trait has a zero heritability. Example: The length of the seeds of a pure strain of beans may vary over several millimeters. However, if extra-large beans are mated, the new crop shows no shift to a larger size. So the heritability of length is zero. On the other hand, if the offspring of two extra-large mice are just as large as they are, genes are probably at work. The trait is said to have a heritability of 1.
The Effects of Selection on Populations
The pressures of natural selection can affect the distribution of phenotypes in a population in several ways.

Stabilizing Selection
Natural selection often works to weed out individuals at both extremes of a range of phenotypes resulting in the reproductive success of those near the mean. In such cases, the result is to maintain the status quo. It is not always easy to see why both extremes should be handicapped; perhaps sexual selection or liability to predation is at work. In any case, stabilizing selection is common. In humans, for example, the incidence of infant mortality is higher for very heavy as well as for very light babies.
Directional Selection
A population may find itself in circumstances where individuals occupying one extreme in the range of phenotypes are favored over the others. Since 1973, Peter and Rosemary Grant - aided by a succession of colleagues - have studied Darwin's finches in the Galapagos Islands. When rainfall, and thus food, are plentiful, the ground finches tend to have a varied diet, e.g., eat seeds of a range of sizes and show considerable variation in body and beak size (large beaks are better for large seeds but can handle small seeds as well as small beaks).
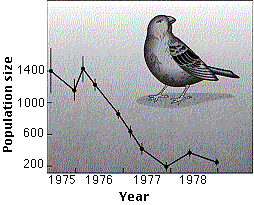
From 1976 through 1977, a severe drought struck the islands, with virtually no rainfall for over a year. This caused a precipitous decline in the production of the seeds that are the dietary mainstay of Geospiza fortis, the medium ground finch. The graph (from P. T. Boag and P. R. Grant in Science 214:82, 1981) shows how the population declined from 1400 to 200 on the island of Daphne Major, a tiny (10-acre = 4 hectares) member of the Galapagos Islands.
One of the plants to make it through the drought produces seeds in large, tough fruits that are virtually impossible for birds with a beak smaller than 10.5 mm to eat. Sampling the birds that died as well as those that survived showed that he larger birds were favored over the smaller ones and Those with larger beaks were favored over those with smaller ones.
| Beak length (mm) | Beak depth (mm) | |
|---|---|---|
| Dead birds | 10.68 | 9.42 |
| Survivors | 11.07 | 9.96 |
Here, then was natural selection at work. But did it produce evolution? The answer turned out to be yes. As the population of G. fortis recovered after the rains returned, the average body size and beak depth of their offspring was greater than before (an increase of 4–5% for beak depth). The bell-shaped curve had been shifted to the right — directional selection.
More recently, the Grants and colleagues at Harvard Medical School have shown that
- beak width and depth in the ground finches are correlated with the timing and intensity of expression of the gene, Bmp4, (that encodes bone morphogenetic protein-4) in the tissue that will form the upper beak. Bmp4 expression appears earlier in development and with greater intensity in the large-beaked Geospiza magnirostris (the large ground finch) than in its smaller-beaked relatives, Geospiza fortis (the medium ground finch) and Geospiza fuliginosa (the small ground finch). See Abzhanov, A., et al., Science, 3 September 2004.
- However, beak length is correlated with the intensity of expression of the gene CaM that encodes the Ca2+-binding protein calmodulin in the tissue that will form the upper beak. CaM expression is much higher in the embryonic tissue of the cactus finches (G. scandens and G. conirostris - both with long beaks) than in their short-beaked relatives, the ground finches G. fortis and G. magnirostris. (See Abzhanov, A., et al., Nature, 3 August 2006.)
Industrial Melanism
Many species of moths in the British Isles began to become darker in color in the 19th century. The best-studied example is the peppered moth, Biston betularia. The moth gets its name from the scattered dark markings on its wings and body. In 1849, a coal-black mutant was found near Manchester, England. Within a century, this black form had increased to 90% of the population in this region. The moth flies at night and rests by day on tree trunks. In areas far from industrial activity, the trunks of trees are encrusted with lichens. As the photos show, the light form (circled in red) is practically invisible against this background.

In areas where air pollution is severe, the combination of toxic gases and soot has killed the lichens and blackened the trunks. Against such a background, the light form stands out sharply. The moth is preyed upon by birds that pluck it from its resting place by day. In polluted woods, the dark form has a much better chance of surviving undetected. When the English geneticist H. B. D. Kettlewell (who supplied the photos) released moths of both types in the woods, he observed that birds did, indeed, eat a much higher fraction of the light moths he released than of the dark. Since pollution abatement programs were put in place after World War II, the light form has been making a comeback in the Liverpool and Manchester areas.
Disruptive Selection
In some circumstances, individuals at both extremes of a range of phenotypes are favored over those in the middle. This is called disruptive selection.
An example:
The residues ("tailings") of mines often contain such high concentrations of toxic metals (e.g., copper, lead) that most plants are unable to grow on them. However, some hardy species (e.g. certain grasses) are able to spread from the surrounding uncontaminated soil onto such waste heaps. These plants develop resistance to the toxic metals while their ability to grow on uncontaminated soil decreases. Because grasses are wind pollinated, breeding between the resistant and nonresistant populations goes on. But evidently, disruptive selection is at work. Higher death rates of both less resistant plants growing on contaminated soil and more resistant plants growing on uncontaminated soil leads to increasing divergence of the populations into two subpopulations with the extreme manifestations of this trait.
The evolutionary significance of disruptive selection lies in the possibility that the gene pool may become split into two distinct gene pools. This may be a way in which new species are formed. The formation of one or more species from a single precursor species is called speciation.