
16.2E: Photorespiration and C4 Plants
- Page ID
- 5786

All plants carry on photosynthesis by adding carbon dioxide (CO2) to a phosphorylated 5-carbon sugar called ribulose bisphosphate. This reaction is catalyzed by the enzyme ribulose bisphosphate carboxylase oxygenase (RUBISCO). The resulting 6-carbon compound breaks down into two molecules of 3-phosphoglyceric acid (PGA). These 3-carbon molecules serve as the starting material for the synthesis of glucose and other food molecules. The process is called the Calvin cycle and the pathway is called the C3 pathway.
Photorespiration
As its name suggests, RUBISCO catalyzes two different reactions:
- adding CO2 to ribulose bisphosphate — the carboxylase activity
- adding O2 to ribulose bisphosphate — the oxygenase activity
Which one predominates depends on the relative concentrations of O2 and CO2 with
- high CO2, low O2 favoring the carboxylase action
- high O2, low CO2 favoring the oxygenase action
The light reactions of photosynthesis liberate oxygen and more oxygen dissolves in the cytosol of the cell at higher temperatures. Therefore, high light intensities and high temperatures (above ~ 30°C) favor the second reaction.
The uptake of O2 by RUBISCO forms the 3-carbon molecule 3-phosphoglyceric acid, just as in the Calvin cycle, and the 2-carbon molecule glycolate. The glycolate enters peroxisomes where it uses O2 to form intermediates that enter mitochondria where they are broken down to CO2. So this process uses O2 and liberates CO2 as cellular respiration does which is why it is called photorespiration. It undoes the good anabolic work of photosynthesis, reducing the net productivity of the plant. For this reason, much effort so far largely unsuccessful has gone into attempts to alter crop plants so that they carry on less photorespiration. The problem may solve itself. If atmospheric CO2 concentrations continue to rise, perhaps this will enhance the net productivity of the world's crops by reducing losses to photorespiration.
C4 Plants
Over 8,000 species of angiosperms have developed adaptations which minimize the losses to photorespiration. They all use a supplementary method of CO2 uptake which forms a 4-carbon molecule instead of the two 3-carbon molecules of the Calvin cycle. Hence these plants are called C4 plants. (Plants that have only the Calvin cycle are thus C3 plants). Some C4 plants - called CAM plants - separate their C3 and C4 cycles by time, while other C4 plants have structural changes in their leaf anatomy so that their C4 and C3 pathways are separated in different parts of the leaf with RUBISCO sequestered where the CO2 level is high; the O2 level low.
After entering through stomata, CO2 diffuses into a mesophyll cell. Being close to the leaf surface, these cells are exposed to high levels of O2, but they have no RUBISCO so cannot start photorespiration (nor the dark reactions of the Calvin cycle).
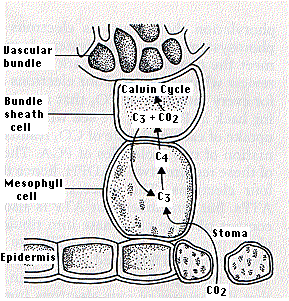
Instead the CO2 is inserted into a 3-carbon compound (C3) called phosphoenolpyruvic acid (PEP) forming the 4-carbon compound oxaloacetic acid (C4). Oxaloacetic acid is converted into malic acid or aspartic acid (both have 4 carbons), which is transported (by plasmodesmata) into a bundle sheath cell. Bundle sheath cells are deep in the leaf so atmospheric oxygen cannot diffuse easily to them and often have thylakoids with reduced photosystem II complexes (the one that produces O2). Both of these features keep oxygen levels low in Bundle sheath cells, which is where the 4-carbon compound is broken down into carbon dioxide, which enters the Calvin cycle to form sugars and starch, and pyruvic acid (C3), which is transported back to a mesophyll cell where it is converted back into PEP.
These C4 plants are well adapted to (and likely to be found in) habitats with high daytime temperatures and intense sunlight. Some examples crabgrass, corn (maize), sugarcane, and sorghum. Although only ~3% of the angiosperms, C4 plants are responsible for ~25% of all the photosynthesis on land.
The ability to use the C4 pathway has evolved repeatedly in different families of angiosperms - a remarkable example of convergent evolution. Perhaps the potential is in all angiosperms.
A report in the 24 January 2002 issue of Nature (by Julian M. Hibbard and W. Paul Quick) describes the discovery that tobacco, a C3 plant, has cells capable of fixing carbon dioxide by the C4 path. These cells are clustered around the veins (containing xylem and phloem) of the stems and also in the petioles of the leaves. In this location, they are far removed from the stomata that could provide atmospheric CO2. Instead, they get their CO2 and/or the 4-carbon malic acid in the sap that has been brought up in the xylem from the roots.
If this turns out to be true of many C3 plants, it would explain why it has been so easy for C4 plants to evolve from C3 ancestors.
CAM Plants
CAM plants are also C4 plants (CAM stands for crassulacean acid metabolism because it was first studied in members of the plant family Crassulaceae.). However, instead of segregating the C4 and C3 pathways in different parts of the leaf, CAM plants separate them in time instead (Table 1).
| Night | Morning |
|---|---|
|
|
These adaptations also enable their owners to thrive in conditions of high daytime temperatures, intense sunlight, and low soil moisture. Some examples of CAM plants include cacti, Bryophyllum, the pineapple and all epiphytic bromeliads, sedums, and the "ice plant" that grows in sandy parts of the scrub forest biome.

C4 Diatoms
On 26 October 2000, Nature reported the discovery of both the C3 and C4 pathways in a marine diatom. In this unicellular organism, the two paths are kept separate by having the C4 path in the cytosol, and the C3 path confined to the chloroplast. The presence of a C4 pathway probably reflects the frequent low concentrations of CO2 in ocean waters.