
15.9C: Vision
- Page ID
- 5686

The human eye is wrapped in three layers of tissue:
- the sclerotic coat. This tough layer creates the "white" of the eye except in the front where it forms the transparent cornea. The corneaThe surface of the cornea is kept moist and dust-free by secretions from the tear glands.
- admits light to the interior of the eye and
- bends the light rays to that they can be brought to a focus.
- the choroid coat. This middle layer is deeply pigmented with melanin. It reduces reflection of stray light within the eye. The choroid coat forms the iris in the front of the eye. This, too, is pigmented and is responsible for eye "color". The size of its opening, the pupil, is variable and under the control of the autonomic nervous system. In dim light (or when danger threatens), the pupil opens wider letting more light into the eye. In bright light the pupil closes down. This not only reduces the amount of light entering the eye but also improves its image-forming ability (as does "stopping down" the iris diaphragm of a camera).
- the retina The retina is the inner layer of the eye. It contains the light receptors, the rods and cones (and thus serves as the "film" of the eye). The retina also has many interneurons that process the signals arising in the rods and cones before passing them back to the brain. (Note: the rods and cones are not at the surface of the retina but lie underneath the layer of interneurons.)
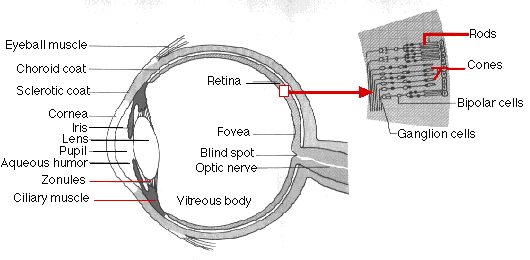
All the nerve impulses generated in the retina travel back to the brain by way of the axons in the optic nerve (above). At the point on the retina where the approximately 1 million axons converge on the optic nerve, there are no rods or cones. This spot, called the blind spot, is thus insensitive to light.

You can demonstrate the presence of the blind spot. Cover your right eye with your hand and stare at the red circle as you move closer to the screen (the square will disappear). Or cover your left eye and stare at the red square as you move.
The Lens
The lens is located just behind the iris. It is held in position by zonules extending from an encircling ring of muscle. When this ciliary muscle is relaxed, its diameter increases, the zonules are put under tension, and the lens is flattened and when contracted, its diameter is reduced, the zonules relax, and the lens becomes more spherical. These changes enable the eye to adjust its focus between far objects and near objects.
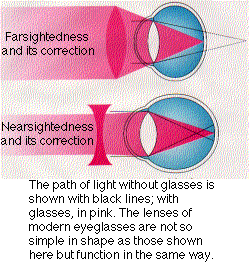
Farsightedness. If the eyeball is too short or the lens too flat or inflexible, the light rays entering the eye — particularly those from nearby objects — will not be brought to a focus by the time they strike the retina. Eyeglasses with convex lenses can correct the problem. Farsightedness is called hypermetropia.
Nearsightedness. If the eyeball is too long or the lens too spherical, the image of distant objects is brought to a focus in front of the retina and is out of focus again before the light strikes the retina. Nearby objects can be seen more easily. Eyeglasses with concave lenses correct this problem by diverging the light rays before they enter the eye. Nearsightedness is called myopia.
Cataracts One or both lenses often become cloudy as one ages. When a cataract seriously interferes with seeing, the cloudy lens is easily removed and a plastic one substituted. The entire process can be done in a few minutes as an outpatient under local anesthesia.
The iris and lens divide the eye into two main chambers:
- the front chamber is filled with a watery liquid, the aqueous humor
- the rear chamber is filled with a jellylike material, the vitreous body
The Retina
Four kinds of light-sensitive receptors are found in the retina:
- rods
- three kinds of cones, each "tuned" to respond best to light from a portion of the spectrum of visible light
- cones that absorb long-wavelength light (red)
- cones that absorb middle-wavelength light (green)
- cones that absorb short-wavelength light (blue)
This scanning electron micrograph (courtesy of Scott Mittman and David R. Copenhagen) shows rods and cones in the retina of the tiger salamander. Each type of receptor has its own special pigment for absorbing light. Each consists of a transmembrane protein called opsin coupled to the prosthetic group retinal. Retinal is a derivative of vitamin A (which explains why night blindness is one sign of vitamin A deficiency) and is used by all four types of receptors.

The amino acid sequence of each of the four types of opsin are similar, but the differences account for their differences in absorption spectrum. The retina also contains a complex array of interneurons:
- bipolar cells and ganglion cells that together form a path from the rods and cones to the brain
- a complex array of other interneurons that form synapses with the bipolar and ganglion cells and modify their activity.
Ganglion cells are always active. Even in the dark they generate trains of action potentials and conduct them back to the brain along the optic nerve. Vision is based on the modulation of these nerve impulses. There is not the direct relationship between visual stimulus and an action potential that is found in the senses of hearing, taste, and smell. In fact, action potentials are not even generated in the rods and cones.
Rod Vision
Rhodopsin is the light-absorbing pigment of the rods. This G-protein-coupled receptor (GPCR) is incorporated in the membranes of disks that are neatly stacked (some 1000 or more of them) in the outer portion of the rod. This arrangement is reminiscent of the organization of thylakoids, another light-absorbing device.
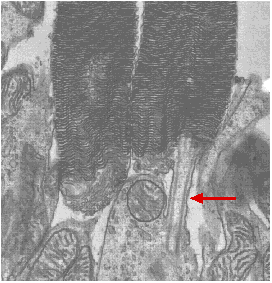
Fig.15.9.3.5 Rod cells of kangaroo bat
The electron micrograph (courtesy of Keith Porter) shows the rod cells of the kangaroo rat. The outer segments of the rods contain the orderly stacks of membranes which incorporate rhodopsin. The inner portion contain many mitochondria. The two parts of the rod are connected by a stalk (arrow) that has the same structure as a primary cilium. Although the disks are initially formed from the plasma membrane, they become separated from it. Thus signals generated in the disks must be transmitted by a chemical mediator (a "second messenger" called cyclic GMP (cGMP) to alter the potential of the plasma membrane of the rod. Rhodopsin consists of an opsin of 348 amino acids coupled to retinal. Like all G-protein-coupled receptors, opsin has 7 segments of alpha helix that pass back and forth through the lipid bilayer of the disk membrane.
Retinal
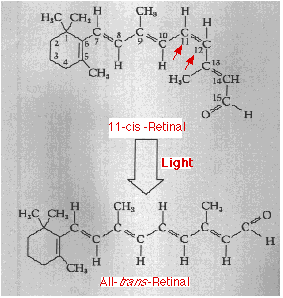
Retinal consists of a system of alternating single and double bonds. In the dark, the hydrogen atoms attached to the #11 and #12 carbon atoms of retinal (red arrows) point in the same direction producing a kink in the molecule. This configuration is designated cis. When light is absorbed by retinal, the molecule straightens out forming the all-trans isomer.
![]()
This physical change in retinal triggers the following chain of events culminating in a change in the pattern of impulses sent back along the optic nerve.
- Formation of all-trans retinal activates its opsin.
- Activated rhodopsin, in turn, activates many molecules of a G protein called transducin.
- Transducin activates an enzyme that breaks down cyclic GMP.
- The drop in cGMP closes Na+ channels in the plasma membrane of the rod. Because these positive ions can no longer enter, the interior of the cell becomes more negative (hyperpolarized) increasing its membrane potential from about −30 to some −70 mV.
- This slows the release of the neurotransmitter glutamate at synapses between the rod and interneurons (e.g., bipolar cells).
- This reduction in glutamate release activates some interneuron pathways, inhibits others.
- The interplay of excited and inhibited interneurons modulates the spontaneous firing of the ganglion cells to which they are connected and gives rise to the ability of the retina to discriminate shapes.
So the retina is not simply a sheet of photocells, but a tiny brain center that carries out complex information processing before sending signals back along the optic nerve. In fact, the retina really is part of the brain and grows out from it during embryonic development.
Rod vision is acute but coarse
Rods do not provide a sharp image for several reasons.
- Adjacent rods are connected by gap junctions and so share their changes in membrane potential.
- Several nearby rods often share a single circuit to one ganglion cell.
- A single rod can send signals to several different ganglion cells.
So if only a single rod is stimulated, the brain has no way of determining exactly where on the retina it was. However, rods are extremely sensitive to light. A single photon (the minimum unit of light) absorbed by a small cluster of adjacent rods is sufficient to send a signal to the brain. So although rods provide us with a relatively grainy, colorless image, they permit us to detect light that is over a billion times dimmer than what we see on a bright sunny day.
Cone Vision
Although cones operate only in relatively bright light, they provide us with our sharpest images and enable us to see colors. Most of the 3 million cones in each retina are confined to a small region just opposite the lens called the fovea. So our sharpest and colorful images are limited to a small area of view. Because we can quickly direct our eyes to anything in view that interests us, we tend not to be aware of just how poor our peripheral vision is.
The three types of cones provide us the basis of color vision. Cones are "tuned" to different portions of the visible spectrum.
- red absorbing cones; those that absorb best at the relatively long wavelengths peaking at 565 nm
- green absorbing cones with a peak absorption at 535 nm
- blue absorbing cones with a peak absorption at 440 nm.
Retinal is the prosthetic group for each pigment. Differences in the amino acid sequence of their opsins accounts for the differences in absorption. The response of cones is not all-or-none. Light of a given wavelength (color), say 500 nm (green), stimulates all three types of cones, but the green-absorbing cones will be stimulated most strongly. Like rods, the absorption of light does not trigger action potentials but modulates the membrane potential of the cones.
Color Blindness
The term color blindness is something of a misnomer. Very few (~1 in 105) people cannot distinguish colors at all. Most "color-blind" people actually have abnormal color vision such as confusing the red and green of traffic lights. As high as 8% of the males in some populations have an inherited defect in their ability to discriminate reds and greens. These defects are found almost exclusively in males because the genes that encode the red-absorbing and green-absorbing opsins are on the X chromosome.
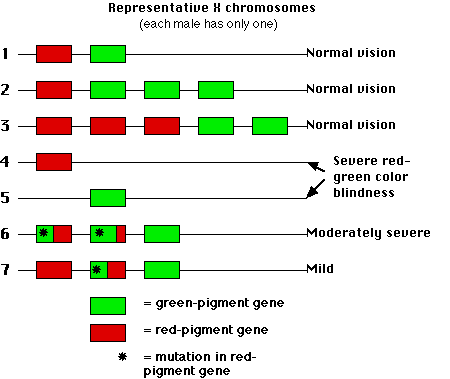
The X chromosome normally carries a cluster of from 2 to 9 opsin genes. The minimum basis for normal red-green vision is one gene whose opsin absorbs efficiently in the red and one that absorbs well in the green (chromosome 1 in the figure). Multiple copies of these genes are also fine (2 and 3). Males with either a "green gene" or "red gene" missing are severely color blind (4 and 5). However, if all the red genes carry mutations (this seldom seems to be the case for the green genes — nobody knows why), then they may have red-green color blindness that ranges from mild to severe depending on the particular mutations involved (6). The rule seems to be that the more the mutations shift the pigment towards green, the more serious the defect. However, a large number of mutations don't always produce serious defects. Multiple mutations in a single gene may offset each other producing only mild defects. And as long as one normal copy of each gene is present, the presence of additional mutated genes seldom produce a serious problem (7).
Why do some males have as many as 9 copies of genes encoding the red and green opsins, when two are enough? The sequences of the red and green genes are the same at 98% of their nucleotides. This high degree of similarity creates the risk of mismatches in synapsis during meiosis with unequal crossing over.
Blue vision
Abnormal blue sensitivity occasionally occurs in humans but is much rarer than abnormalities in red-green vision. The gene for the blue-cone opsin is located on chromosome 7. Thus this trait shows an autosomal pattern of inheritance being found in females as often as in males.