
15.4C: B Cells and T Cells
- Page ID
- 5432

Lymphocytes are one of the five kinds of white blood cells (leukocytes) that circulate in the blood. Although mature lymphocytes all look pretty much alike, they are extraordinarily diverse in their functions. The most abundant lymphocytes are B lymphocytes (B cells) and T lymphocytes (T cells). B cells are produced in the bone marrow. The precursors of T cells are also produced in the bone marrow but leave the bone marrow and mature in the thymus (which accounts for their designation). Each B cell and T cell is specific for a particular antigen. What this means is that each is able to bind to a particular molecular structure. The specificity of binding resides in a receptor for antigen: the B cell receptor (BCR) for antigen and the T cell receptor (TCR) respectively.
Both BCRs and TCRs share these properties:
- They are integral membrane proteins.
- They are present in thousands of identical copies exposed at the cell surface.
- They are made before the cell ever encounters an antigen.
- They are encoded by genes assembled by the recombination of segments of DNA.
- They have a unique binding site.
- This site binds to a portion of the antigen called an antigenic determinant or epitope.
- The binding, like that between an enzyme and its substrate depends on complementarity of the surface of the receptor and the surface of the epitope.
- The binding occurs by non-covalent forces (again, like an enzyme binding to its substrate).
- Successful binding of the antigen receptor to the epitope, if accompanied by additional signals, results in:
- stimulation of the cell to leave G0 and enter the cell cycle.
- Repeated mitosis leads to the development of a clone of cells bearing the same antigen receptor; that is, a clone of cells of the identical specificity.
BCRs and TCRs differ in their structure, the genes that encode them and the type of epitope to which they bind.
B Cells
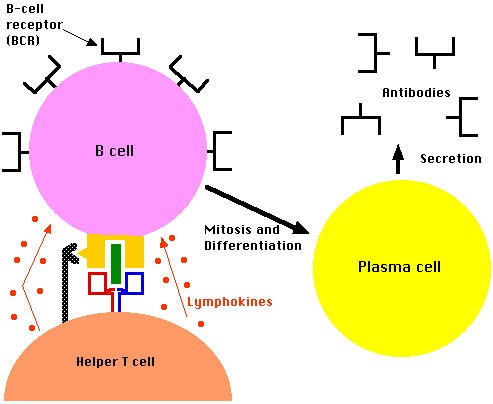
Fig.15.4.3.1 B cell and lymphokines
- BCRs bind intact antigens (like diphtheria toxoid, the protein introduced into your body in the DTP vaccine). These may be
- soluble molecules present in the extracellular fluid
- intact molecules that the B cell plucks from the surface of antigen-presenting cells like macrophages and dendritic cells
- The bound antigen molecules are engulfed into the B cell by receptor-mediated endocytosis.
- The antigen is digested into fragments which are then displayed at the cell surface nestled inside a class II histocompatibility molecule.
- Helper T cells specific for this structure (i.e., with complementary TCRs) bind the B cell and secrete lymphokines that:
- stimulate the B cell to enter the cell cycle and develop, by repeated mitosis, into a clone of cells with identical BCRs
- switch from synthesizing their BCRs as integral membrane proteins to a soluble version
- differentiate into plasma cells that secrete these soluble BCRs, which we now call antibodies
T Cells
The surface of each T cell also displays thousands of identical T cell receptors (TCRs). There are two types of T cells that differ in their TCR:
- alpha/beta (αβ) T cells. Their TCR is a heterodimer of an alpha chain with a beta chain. Each chain has a variable (V) region and a constant (C) region. The V regions each contain 3 hypervariable regions that make up the antigen-binding site.
- gamma/delta (γδ) T cells. Their TCR is also a heterodimer of a gamma chain paired with a delta chain.
The discussion that follows now concerns alpha/beta T cells. Gamma/delta T cells, which are less well understood, are discussed at the end.
The TCR (of alpha/beta T cells) binds a bimolecular complex displayed at the surface of some other cell called an antigen-presenting cell (APC). This complex consists of a fragment of an antigen lying within the groove of a histocompatibility molecule. The complex has been compared to a "hot dog in a bun".

Most of the T cells in the body belong to one of two subsets. These are distinguished by the presence on their surface of one or the other of two glycoproteins designated:
- CD4
- CD8
Which of these molecules is present determines what types of cells the T cell can bind to.
- CD8+ T cells bind epitopes that are part of class I histocompatibility molecules. Almost all the cells of the body express class I molecules.
- CD4+ T cells bind epitopes that are part of class II histocompatibility molecules. Only specialized antigen-presenting cells express class II molecules. These include dendritic cells, phagocytic cells like macrophages and B cells.
CD8+ T cells
The best understood CD8+ T cells are cytotoxic T lymphocytes (CTLs). They secrete molecules that destroy the cell to which they have bound.

This is a very useful function if the target cell is infected with a virus because the cell is usually destroyed before it can release a fresh crop of viruses able to infect other cells. An example will show the beauty and biological efficiency of this mechanism.
Every time you get a virus infection, say influenza (flu), the virus invades certain cells of your body (in this case cells of the respiratory passages). Once inside, the virus subverts the metabolism of the cell to make more virus. This involves synthesizing a number of different macromolecules encoded by the viral genome. In due course, these are assembled into a fresh crop of virus particles that leave the cell (often killing it in the process) and spread to new target cells. Except while in transit from their old homes to their new, the viruses work inside of your cells safe from any antibodies that might be present in blood, lymph, and secretions. But early in the process, infected cells display fragments of the viral proteins in their surface class I molecules. CTLs specific for that antigen will be able to bind to the infected cell and often will be able to destroy it before it can release a fresh crop of viruses.
In general, the role of the CD8+ T cells is to monitor all the cells of the body, ready to destroy any that express foreign antigen fragments in their class I molecules.
CD4+ T cells
CD4+ T cells bind an epitope consisting of an antigen fragment lying in the groove of a class II histocompatibility molecule. CD4+ T cells are essential for both the cell-mediated and antibody-mediated branches of the immune system:
- cell-mediated immunity: These CD4+ cells bind to antigen presented by antigen-presenting cells (APCs) like phagocytic macrophages and dendritic cells. The T cells then release lymphokines that attract other cells to the area. The result is inflammation: the accumulation of cells and molecules that attempt to wall off and destroy the antigenic material (an abscess is one example, the rash following exposure to poison ivy is another).
- antibody-mediated immunity: These CD4+ cells, called helper T cells, bind to antigen presented by B cells. The result is the development of clones of plasma cells secreting antibodies against the antigenic material.
AIDS provides a vivid illustration of the importance of CD4+ T cells in immunity. The human immunodeficiency virus (HIV) binds to CD4 molecules and thus is able to invade and infect CD4+ T cells. As the disease progresses, the number of CD4+ T cells declines below its normal level of about 1000 per microliter (µl). A partial explanation for this may be the unceasing efforts of the patient's CD8+ T cells to destroy the infected CD4+ cells. However, it turns out that only a small fraction of the patients CD4+ T cells are infected at any given time. How uninfected CD4+ cells may be induced to commit suicide is discussed in the page on apoptosis.
When the number of CD4+ T cells drops below 400 per microliter, the ability of the patient to mount an immune response declines dangerously. Not only does the patient become hypersusceptible to pathogens that give all of us grief but also to microorganisms, especially viruses and fungi, that normally inhabit our tissues without harming us. These opportunistic infections can be fatal.
Building the T-cell Repertoire
T cells have receptors (TCRs) that bind to antigen fragments nestled in MHC molecules. But all cells express class I MHC molecules containing fragments derived from self proteins. Many cells express class II MHC molecules that also contain self peptides. This presents a risk to the animal of the T cells recognizing these self-peptide/self-MHC complexes and mounting an autoimmune attack against them. Fortunately, this is usually avoided by a process of selection that goes on in the thymus (where all T cells develop).
The process works like this:
- The precursors of T cells like all blood cells are formed in the bone marrow.
- These cells then migrate to the cortex of the thymus. At this time they have neither a complete TCR nor either CD4 or CD8 (thus are called "double-negative" or DN cells).
- In the cortex of the thymus they begin to form a TCR and synthesize both CD4 and CD8 (so now they are "double-positive" or DP cells). The cortical cells of the thymus express a wide variety of small molecules, usually a peptide of 6–8 amino acids derived from body proteins; that is, "self" proteins such as proteins within the cytosol and serum proteins - proteins circulating in the blood and lymph nestled in a histocompatibility molecule (encoded by the MHC).
- Most of the cells (~97%) will produce a TCR that does not bind to any of the peptide-MHC molecules present on the surface of the cortical cells. Unless they can try again with a new TCR, these cells die by "neglect" (by apoptosis, actually).
- Those remaining cells whose TCR has bound a peptide antigen presented in class II MHC molecule stop expressing CD8 and become CD4+ T cells. It is these cells that will go on to become
- Th1 cells in cell-mediated immune responses
- Th1 helper cells for cytotoxic T lymphocytes (CTLs)
- Th2 helper cells for B cells
- Those remaining cells whose TCR has bound a peptide antigen presented in class I MHC molecule stop expressing CD4 and become CD8+ T cells.
- Both sets of cells are said to have undergone positive selection.
- After positive selection, these cells migrate to the medulla of the thymus.
- There those cells whose TCR binds very strongly to complexes of self-peptide and self-MHC are destroyed (again by apoptosis).
- This process of negative selection is important as it eliminates cells that might otherwise mount an autoimmune attack. It is one of the ways in which tolerance to self antigens is achieved. [Link to discussion of T-cell tolerance.]
- The cells whose TCRs bind antigen at an affinity below the threshold that triggers apoptosis are free to leave the thymus and migrate throughout the immune system (lymph nodes, spleen, etc.)
- It is this population that we depend on to mount immune responses against foreign antigens. A TCR that binds self-peptide/self-MHC with low affinity may well bind a foreign-peptide in self MHC with high affinity.
Gamma/Delta (γδ) T Cells
Gamma/delta T cells differ from their alpha/beta cousins in several ways:
- Their TCR is encoded by different gene segments.
- Their TCR binds to
- antigens that can be intact proteins (just as antibodies do) as well as a variety of other types of organic molecules (often containing phosphorus atoms).
- antigens that are not "presented" within class I or class II histocompatibility molecules;
- antigens that are not presented by "professional" antigen-presenting cells (APCs) like dendritic cells.
- Most of these T cells have neither CD8 nor CD4 on their surface. This makes sense because they have no need to recognize class I and class II histocompatibility molecules.
- Gamma/Delta T cells, like alpha/beta T cells, develop in the thymus. However, they migrate from there into body tissues, especially epithelia (e.g., intestine, skin, lining of the vagina), and don't recirculate between blood and lymph nodes (they represent no more than 5% of the T cells in the blood and are even rarer in lymph nodes). They encounter antigens on the surface of the epithelial cells that surround them rather than relying on the APCs found in lymph nodes.
What is the Function of γδ T cells?
That is still something of a mystery. Situated as they are at the interfaces between the external and internal worlds, they may represent a first line of defense against invading pathogens. Their response does seem to be quicker than that of αβ T cells. Curiously, many of the antigens to which γδ T cells respond are found not only on certain types of invaders (e.g., Mycobacterium tuberculosis, the agent of tuberculosis) but also on host cells that are under attack by pathogens.
Knockout mice that cannot make γδ T cells are slower to heal injuries to their skin. They are also much more susceptible to skin cancers than normal mice. Perhaps immune surveillance is one of the functions of γδ T cells.
Contributors and Attributions
John W. Kimball. This content is distributed under a Creative Commons Attribution 3.0 Unported (CC BY 3.0) license and made possible by funding from The Saylor Foundation.