
10.1: Case Study - Ectodermal Appendages
- Page ID
- 20774

Ectodermal appendages are organs associated with the skin like feathers and hair. We typically only call vertebrate skin elaborations "ectodermal appendages" even though other animal groups have skin organs (like glands) as well. In this section we will just focus on the vertebrate appendages with a special focus on the mammary glands. Details on this as well as a summary of the development of other skin organs can be found here: Mechanisms of ectodermal organogenesis.
Ectodermal appendages (or organs) have common developmental patterning and are thought to have a common evolutionary organ as well, with co-option of elements from different appendage types occurring frequently. These appendages are first obvious as a thickening of the epidermis to form a "placode." This is followed by cellular proliferation of the epidermis and condensation of underlying mesenchymal cells of the dermis to form a bud with a thick mesenchymal base. The ectodermally derived bud can now undergo one of multiple trajectories, which it chooses based on signals from the mesoderm - the mesenchyme of the dermis as well as the somitic mesoderm (Figure 1). We roughly classify these appendages into two main groups: the oral appendages, including teeth and salivary glands, and the skin appendages, including feathers, scales, hair, mammary glands, sweat glands, and oil glands.

Early on, scientists noticed something interesting - once an ectodermal placode was in place, it could be induced to form one of many tissues. For example, mammary bud mesenchyme can stimulate the formation of mammary glands in dorsal epithelium2. One interesting experiment showed that mouse dental mesenchyme could even induce teeth in chick oral epithelium, despite the fact that birds do not ordinarily grow teeth3. As discussed earlier, ectodermal appendage development follows the trajectory of general to specific (Figure 2) with the early placodes being less specified than later buds.

Evolution and Diversity of Tetrapod Skin Appendages
Fishes have their own amazing and diverse skin appendages including the "lateral line" organs that detect electrical fields, pressure and water flow changes4. However, here we will focus on the ectodermal appendages of the tetrapods with a focus on the mammary glands of Mammalia. The ancestral tetrapod is hypothesized to have had mucus glands in its skin, similar to those in living amphibians. These mucus glands keep the skin moist and prevent the internal parts of the animal from drying out. Additionally, mucus glands and other skin glands are used by brooding modern amphibians to keep their eggshell-less eggs from drying out in terrestrial ecosystems and may have played a similar role in early tetrapods (Figure 3).

About 350 million years ago, one group of tetrapods, the Amniota, evolved a new type of skin protein that prevented their skin from drying out: keratin. All amniotes (including mammals, birds, lizards, etc) have alpha-keratin and the sauropsids (birds and other reptiles) additionally have beta-keratin, which they use to build feathers and scales. Unlike egg-laying mammals, sauropsids have calcified eggshells which reduce water loss. The egg-laying mammals (the monotremes like platypus), have parchment shelled eggs that can dry out quickly. This is thought to be the ancestral condition for the group containing the mammals, the synapsids, and glandular skin secretions from the brooding parent (like mucus secretions in salamanders) are thought to have helped prevent egg desiccation in ancestral synapsids including mammals5.
The ancestral mammal thus inherited glandular skin that was multifunctional: it likely secreted oil, sweat, and other dilute secretions (possibly a form of sweat) to keep it's eggs hydrated. We can get some hints as to how these skin glands diversified into multiple ectodermal appendages by comparing the three living clades of mammmals the egg-laying monotremes, the therian marsupials, and the therian placentals (also known as the eutherians). Mammals have four main types of skin glands, these are the wax producing glands of the ear (which we will ignore for now, the sebaceous glands that produce oil, the sweat glands, and the mammary glands. The last three of these glands have special anatomical relationships with each other. A typical hair follicle is associated with a sebaceous gland and can also be associated with an apocrine sweat gland. The mammary glands in some species are associated with a hair follicle as well as a sebaceous gland. This has led to the hypothesis that mammary glands are at least partly derived from apocrine sweat glands (Figure 4).
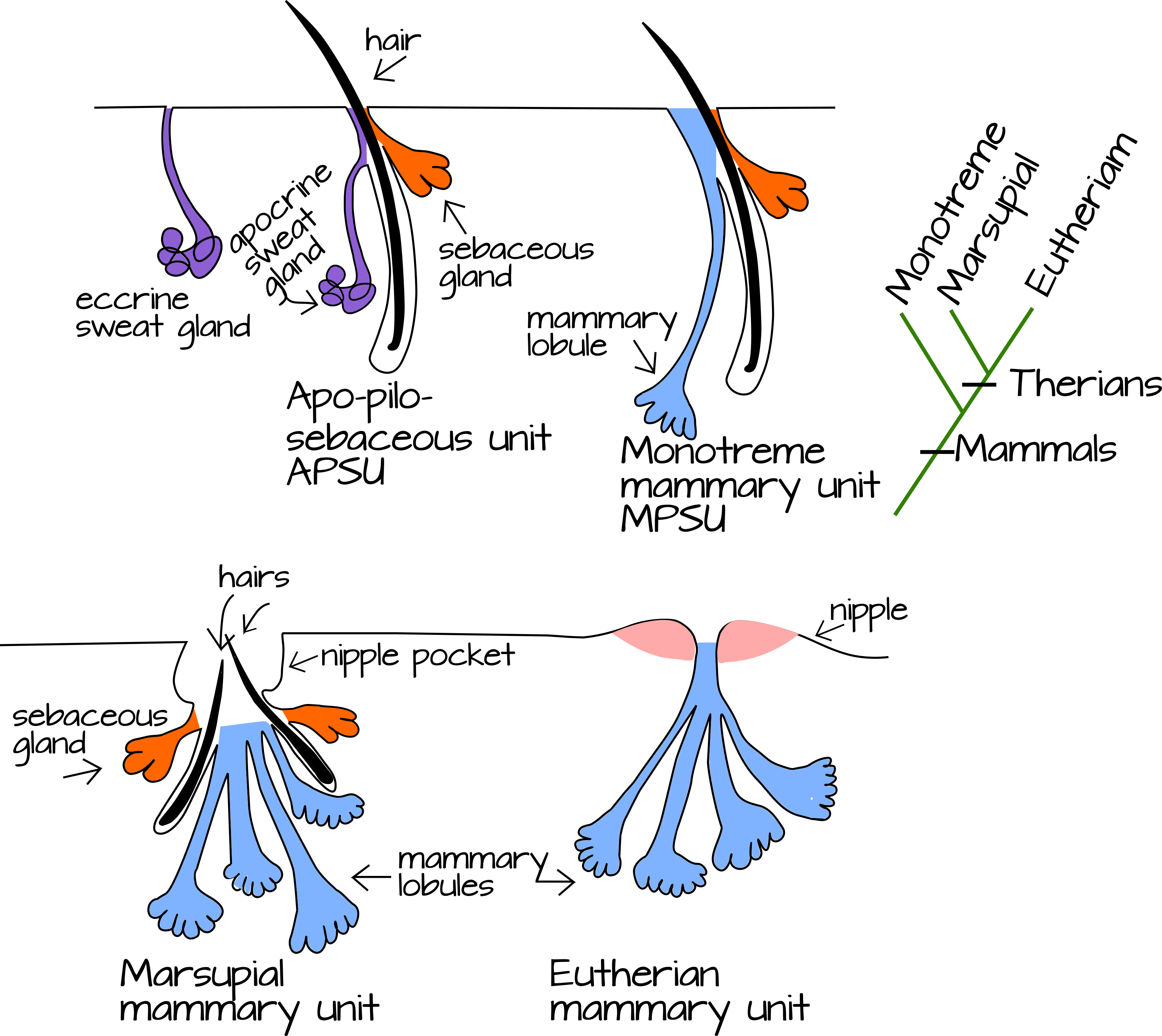 |
Figure 4: Comparison of mammalian skin glands. The APSU (apo-pilosebaceous unit) is a common type of skin gland found in the armpit and groin of humans. It consists of a sebaceous gland, a hair follicle, and an apocrine sweat gland. Apocrine glands secrete by pinching off part of a secretory cell and releasing it into the duct. This is very different from typical (eccrine) sweat gland that secrete by exocytosis and are found all over the skin of humans. The MPSU (mammolobular-pilosebaceous unit) is thought to be a derived APSU with apocrine mammolobular secretory cells replacing the sweat-gland cells. A mammary gland contains one or more MPSUs and a nipple (in most cases, though some mammals like monotremes lack nipples). |
Overview of mammary gland development
Despite the diversity in their final forms, mammary glands develop along largely similar pathways across mammals. As shown in Figure 1, they are induced by underlying mesenchyme and other signals to form two parallel mammary lines, which are likely induced by the Gli3-induced secretion of FGF10 by a subset of the somitic mesoderm, driving the expression of Wnt10b in the overlying ectoderm and specifying mammary cell fate6. Monotremes retain mammary lines as patches of milk-secreting cells. Cross-talk between the epidermis and dermis (mesenchymal) generate a mammary bud, which grows downward into the mesenchyme to form a mammary sprout. In therians, the mammary buds and sprouts receive BMP4 signaling from the underlying mesenchyme to continue to prevent them from taking the default hair-follicle fate6. In other parts of the skin, Noggin and Shh inhibit BMP4 and allow the default hair fate6. The epithelial mammary buds express PTHrP (Parathyroid Hormone related Peptide), which activates its receptor in the mesenchyme to induce condensation and differentiation of the growing mammary primary sprout. Thus there is cross-talk between the epidermis and dermis (mesenchyme) to generate the mammary bud and induce growth into the primary sprout. In some placental mammals like mice the primary sprout grows deeply into hypodermal fat tissue where it begins to branch into alveolar glandular tissue. In other mammals it branches more shallowly into secondary sprouts, which themselves branch into alveolar glandular tissue8.
Where did the hair go?
If mammary glands evolved from hair follicles and their associated glands, what happened to the hair? While many placental mammals have aereolar hair (Figure 5), most do not have hair associated with the part of the nipple that secretes milk. Additionally, no placental mammals examined develop a hair follicle during or before formation of a mammary sprout. I will get back to the placental mammal issue in a moment, but first I'd like to spend a little time looking at our hairy-aerola cousins.
Monotremes, those basally branching mammals, do maintain hair association with mammary glands (Figure 4). Oftedal proposes that this is the ancestral mammalian case and that mammary hairs, rather than a nipple, have a dual use: they not only wick milk from skin glands into the mouth of an infant, but they also wick moist secretions onto dessication-prone eggs. The evolution of internal embryonic development and live-birth in the therian mammals changed the evolutionary pressure on the milk-producing glands. They lost the selective pressure to keep eggs hydrated and instead were selected on solely for nourishment of infants. This may have driven the milk ducts to condense and flow into a single nipple per mammary gland, rather than a dispersed hair patch (Figure 6)5,8.
Evidence from mammary gland development in therian mammals lends some support to this hypothesis. In marsupials, early mammary gland development proceeds as a physically condensed version of monotreme development. In monotremes, a single mammary primary sprout develops into a hair follicle, a sebaceous gland, and mammary secretory and ductal cells (the multi-component MPSU). In marsupials, a single primary sprout forms multiple MPSUs, each with its own hair follicle, sebaceous gland, and mammary cells. However, the hair follicles eventually atrophy and die, leaving behind a large pocket into which milk and sebum can empty (Figure 4)8. Placental mammals (eutherians), like us, are more variable. Eutherians can make one or many primary sprouts per mammary bulb and (as mentioned above) may make or lack secondary sprouts. Species without secondary sprouts do not have hair follicles and sebaceous associated with growing mammary glands, but species with secondary sprouts do. This includes humans and horses. Our MPSU hair follicles and sebaceous glands atrophy and die during embryogenesis so we are left with an MPSU made of a single component: the mammary alveoli and ducts8.
By comparing the development of the mammary glands of these different groups, Oftedal and Dhouailly have proposed a scenario for mammary gland evolution by co-option and heterochrony. Briefly, mammary glands use existing skin placode developmental mechanisms to begin to form, in particular the existing patterning used by the apo-pilosebaceous glands, then switch to a program of branching morphogenesis (a developmental patterning mechanism found in the kidney, lungs, and salivary glands in tetrapods but also in bird feathers and insect tracheal development). This creates the ductal tree of the mammary gland as the apocrine portion of the apo-pilosebaceous unit, instead of a sweat gland8. The origin of the overlying nipple is still unclear but is developmentally induced by the developing mammary gland.
The genetics of mammary gland evolution: co-option and novelty
Earlier, I briefly discussed the fate choice that skin makes between hair and mammary gland - turning on BMP4 signaling in mesenchymal cells via FGF10 signaling from part of the somitic mesoderm turns off the default hair program and turns on the mammary program. FGF/Wnt/BMP interactions are used in forming other skin in overlying ectoderm is used in patterning other skin appendages and may be a general cassette for ectodermal patterning9. In this way, mammary glands placode and bud formation are likely driven by similar signaling as other ectodermal appendages with some modifications. Once the mammary placode/bud begins to grow, it expresses a common epidermal organ gene: PTHrP. PTHrP has a more ancestral function in vertebrates in endochondrial bone formation and tooth eruption. As teeth are an ectodermal epithelial structure, it is likely that PTHrP's role in mammary gland development was coopted from tooth development. Evidence for co-option of tooth genes comes from another gene family key in tooth development -the calcium binding protein ODAM. ODAM was duplicated during tetrapod evolution to form the caseins, the calcium transport proteins in milk10.
FGF10 and Wnt10b are not the only players in mammary specification. Several Hox genes are implicated in ectodermal organ specification, with Hoxc8 initiating mouse mammary placode development and Hoxb3, b6, b9, d9, d10, and d8, among others, expressed during later development, including during postnatal mammary development11,12. Of these, Hoxd9 and d8 are most commonly expressed in mouse mammary bulbs. These two genes are also involved in both limb and hair development in mice.
While Hoxd9 and d8 are located next to each other on the chromosome and largely regulated by two giant cis-regulatory elements HoxD-T-DOM and HoxD-C-DOM, they do not exhibit completely identical patterns of expression, suggesting that there are specific regulators that act differentialy on their individual promoters. Recently, scientists in the Beccari lab in Geneva dissected the regulatory elements driving Hox d8 and d9 expression and found how they managed to go from being expressed in the limb and hair to being expressed in the mammary bulb. While Hoxd8 mammary bulb expression depended on a 13kb local enhancer region, the Hoxd9 mammary bulb expression depended on distant cis regulatory regions in T-DOM12.
The Hoxd9 mammary bulb cis regulatory element is particularly intriguing because it seems to partly rely on the existing cis regulatory elements that drive limb expression of Hoxd9. Limb expression is driven by cis regulatory elements within T-DOM and during limb development, a giant 1500 kb region of the chromosome folds over to contact Hoxd9. The mammary bud enhancer for Hoxd9 is also found within this 1500 kb region, and it is thought that this enhancer takes advantage of the attraction between the limb bud enhancer and the Hox cluster to get access to Hoxd9 during mammary bulb development (Figure 7)12. Taken together, this begins to paint a picture of the evolution of mammary placodes and bulbs - they use existing transcription factor and patterning genes in new combinations. At least one of the genes involved gets expressed in the developing mammary tissue by co-opting and modifying existing enhancer elements.
Once the embryonic mammary glands are built, they still need to secrete milk. While the similarities between milk-producing glands and apocrine sweat glands are obvious at anatomical and developmental levels, the content of the two substances produced is substantially different. One milk product, the caseins, is made from mutated ODAM tooth protein (see above). The major protein essential in processing the fat droplets found in milk, XOR, is usually an antimicrobial protein but plays a second role in milk biosynthesis. This is a clear case of heterotopy (expression of a gene in a new domain). Lysozyme took both paths - it was coopted from the immune system as an antimicrobial protein in the mammary glands, and it also underwent a gene duplication to form a whey protein used to lactose13. Even though milk composition is complex and varies from species to species (even within species as an infant gets older), we can trace the evolutionary history of these components and see how evolution could have acted to produce this secretion.
References
- "Stem cell fate determination during development and regeneration of ectodermal organs." (2012) Frontiers in Physiology, Lucía Jiménez-Rojo, Zoraide Granchi, Daniel Graf and Thimios A. Mitsiadis, https://doi.org/10.3389/fphys.2012.00107.
- "Mammary Phenotypic Expression Induced in Epidermal Cells by Embryonic Mammary Mesenchyme" (1995) Gerald R. Cunha, Peter Young, Konstantin T Christov, R C Guzman, S. B. Nandi, Frank J. Talamantes, Gudmundur Thordarson, PMID: 7572029.
- "Development of teeth in chick embryos after mouse neural crest transplantations", (2003), PNAS, Thimios A. Mitsiadis, Yvonnick Chéraud, Paul Sharpe, and Josiane Fontaine-Pérus, https://doi.org/10.1073/pnas.1137104100.
- "The lateral line can mediate rheotaxis in fish" (1997), Nature, John C. Montgomery, Cindy F. Baker, and Alexander G. Carton, https://doi.org/10.1038/40135.
- "The Origin of Lactation as a Water Source for Parchment-Shelled Eggs" (2002) Journal of Mammary Gland Biology and Neoplasia, Olav T. Oftedal, https://doi.org/10.1023/A:1022848632125..
- "A new scenario for the evolutionary origin of hair, feather, and avian scales", (2009) Journal of Anatomy, Danielle Dhouailly, doi: 10.1111/j.1469-7580.2008.01041.x.
- "Parathyroid hormone-related protein maintains mammary epithelial fate and triggers nipple skin differentiation during embryonic breast development." (2001) Development, J. Foley, P. Dann, J. Hong, J. Cosgrove, B. Dreyer, D. Rimm, M. Dunbar, W. Philbrick, J. Wysolmerski, PMID: 11171335.
- "Evo-devo of the mammary gland," (2013), Journal of Mammary Gland Biology and Neoplasia, Olav T. Oftedal and Danielle Dhouailly, doi: 10.1007/s10911-013-9290-8.
- "FGF signaling is required for initiation of feather placode development" (2004) Development, Markus Mandler, Annette Neubüser, doi: 10.1242/dev.01203.
- "The evolution of milk casein genes from tooth genes before the origin of mammals" (2012), Kazuhiko Kawasaki, Anne-Gaelle Lafont, Jean-Yves Sire, Molecular Biology and Evolution, https://doi.org/10.1093/molbev/msr020.
- "HOXC8 initiates an ectopic mammary program by regulating Fgf10 and Tbx3 expression, and Wnt/β-catenin signaling" (2015) Development, Lara S. Carroll, Mario R. Capecchi, doi: 10.1242/dev.128298.
- "Control of Hoxd gene transcription in the mammary bud by hijacking a preexisting regulatory landscape," (2016), PNAS, Ruben Schep, Anamaria Necsulea, Eddie Rodríguez-Carballo, Isabel Guerreiro, Guillaume Andrey, Thi Hanh Nguyen Huynh, Virginie Marcet, Jozsef Zákány, Denis Duboule, and Leonardo Beccari, https://doi.org/10.1073/pnas.1617141113.
- "Evolution of the mammary gland from the innate immune system?" (2006) Bioessays, Claudia Vorbach Mario R. Capecchi Josef M. Penninger, doi.org/10.1002/bies.20423.
Thumbnail: Closeup image of peacock feathers, which is one example of Ectodermal Appendages. (Public Domain).