
1.2: EvoDevo Case Study
- Page ID
- 14663

When we look within a species we see tall and short (or large and small) individuals but they are all scaled more or less the same. For example, we don't generally see tall people with abnormally long necks or short people with abnormally short legs. As a general concept, we can say that this kind of scaling must be the result of cross-talk between different cells, tissues, and organs. For example, we might expect a growing bone to stretch muscle fibers and induce them to elongate. We might also expect a bone that grows long to also grow wide in proportion. However, the actual cellular and molecular mechanisms that perform this scaling are often somewhat mysterious. Here we are going to begin to examine a case of scaling in a group of super-manipulable animals - the dipterans.
Bicoid in dipterans: Drosophila
Dipterans are a type of fly, and this group includes the flies you are probably most familiar with - houseflies and Drosophila fruit flies. As was mentioned in the Introduction, these flies undergo extremely rapid development. This is likely selected for by their transient food sources. Moreover, their embryos and larvae lack protection and are a good source of nutrition for insectivorous predators.
One way to speed up development is to pattern the whole animal at the same time instead of patterning one end first, like we and many other animals do. Drosophila and relatives (often called the "higher dipterans") pattern their anterior and posterior ends at the same time. In fact, the oocyte (egg) is patterned before fertilization even occurs, with the "organizer" of anterior patterning (Bicoid) at one end and the "organizer" of posterior patterning (Nanos) at the other1,2. There are many more genes involved in this process, but we will mostly focus here on Bicoid since it has been the subject of intense study.
Both the Nanos and Bicoid organizers in Drosophila are proteins that are found in a concentration gradient. Bicoid has its highest concentration at the anterior end, while Nanos is highest at the posterior. Towards the middle of the embryo they are both present at only low concentrations (Figure 2)1,2,3,4 If you deplete Bicoid from a fly egg, you end up with a fly missing anterior (head) structures and an expanded abdomen. If you deplete Nanos, you end up with a fly missing posterior (abdominal) structures and an expanded head. Note that the tip of the head and the tail are patterned by two separate developmental mechanisms, but we are going to ignore this for now.

At this point, I am going to make our first foray into light molecular genetics. If you get a bit confused, please read Briefly, Genetics for some clarification.
When Christiane Nusslein-Volhard first made Bicoid mutants, she noticed a couple of interesting things. First, you need to make the moms mutant. This makes sense because it is maternal mRNA that gets pumped into the oocyte, so of course it is the mom's genotype that rules the embryo's morphology in this case. Second, even though she didn't have perfect evidence, she noticed that the Bicoid protein gradient looks like a diffusion gradient. A diffusion gradient is the pattern you get when something is diffusing freely from a point source (Figures 2,3,4). Imagine you are baking cookies, a visitor knocks on your door. As you open the door, they notice a faint but terrific smell coming from your house. They make small talk with you and slowly walk towards the source of the smell towards the kitchen. As they get closer to the oven the smell intensifies. You open the oven to take out the cookies, and your kitchen is flooded with the smell of cookie goodness. In this case the baking cookies in your oven are the point source and the smell has formed a diffusion gradient in your house, faintest at the front door and strongest in the kitchen. This mechanism for forming a gradient is called the "SSD model" for synthesis, solute, diffusion. However, in the case of Bicoid, this model has its critics, as we will discover in The Curious Case of Bicoid.

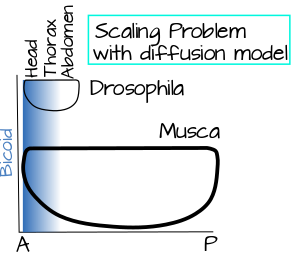
The Problem of Scaling in Dipterans
The SSD model looks good on paper, but there is a confounding fact about dipteran embryos that forces us to take a closer look at it. Dipteran embryos vary in size tremendously. Drosophila embryos are teensy-tiny, about a third to half a centimeter long. Housefly embryos, on the other hand, are a whopping 1.5 centimeters long. What does this mean for the SSD model? Imagine your point source in our cookie example: a delicious smell coming from your oven. Only now you are cooking in your University's dining hall. Someone walks into the dining hall, what is the likelihood that they are going to smell those cookies? Pretty low, unless they have a great sense of smell. The door is now way too far for detectable odor to reach by diffusion. Likewise, in a large Musca (housefly) embryo, the diffusion gradient would only reach 1/5 (or less!) as far down the embryo as the same diffusion gradient in a small Drosophila embryo (Figure 4). How do flies solve this problem? There are several possible solutions that we will consider in The Curious Case of Bicoid.
An editable svg file of the figure above can be downloaded at https://scholarlycommons.pacific.edu/open-images/5/
References
- "The bicoid protein determines position in the Drosophila embryo in a concentration-dependent manner" 1988, Cell, Wolfgang Driever and Christiane Nüsslein-Volhard https://doi.org/10.1016/0092-8674(88)90183-3
- "Nanos is the localized posterior determinant in Drosophila", 1991, Cell, Charlotte Wang and Ruth Lehmann https://doi.org/10.1016/0092-8674(91)90110-K
- "From gradients to stripes in Drosophila embryogenesis: filling in the gaps", 1996, Trends in Genetics, Rolando Rivera-Pomar and Herbert Jãckle https://doi.org/10.1016/0168-9525(96)10044-5
- "Bicoid gradient formation and function in the Drosophila pre-syncytial blastoderm". 2016, eLife, Zehra Ali-Murthy and Thomas B Kornberg, 10.7554/eLife.13222