3.3: The EvoDevo of the Blastopore: The Spemann-Mangold Organizer - Gastrulation in Evolution
- Page ID
- 21530

The Spemann-Mangold Organizer
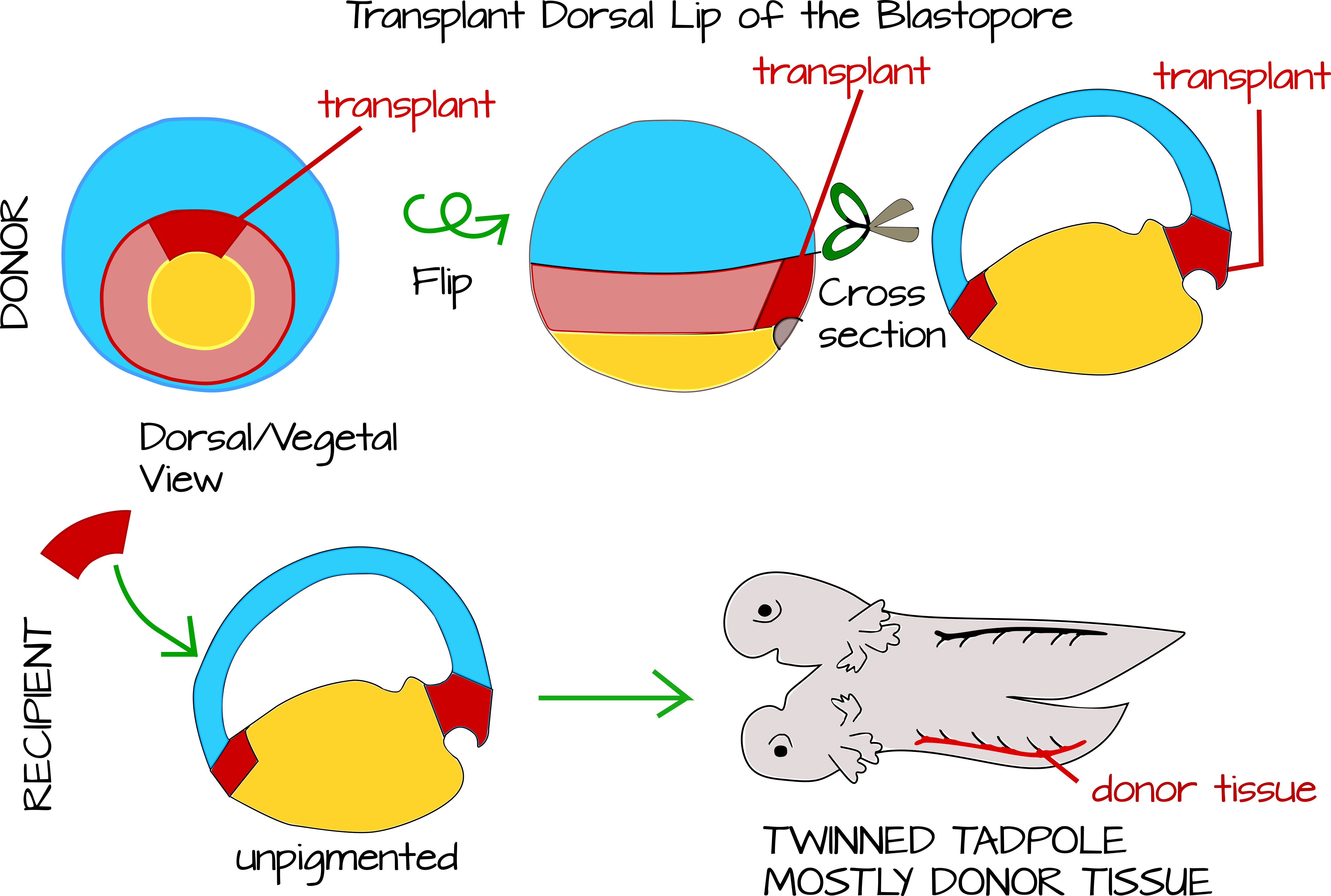
Hans Spemann and his graduate student Hilde Mangold perfected a technique to do cross species transplantations of the dorsal blastopore lip to new locations in the host's body. Spemann had wondered for years what exactly the dorsal lip of the blastopore did. He knew it was important because if he sliced an embryo in two, the half that got the dorsal blastopore lip developed into an tadpole, while the other half developed into a "belly piece." He also knew that if he carefully divided the embryo down the middle of the dorsal lip of the blastopore, both halves would develop into a tadpole. One possibility was that a morphogenic signal emanated from the dorsal lip that caused cellular differentiation in a time gradient, such that the closest cells to the dorsal lip differentiate first. Without this signal, cells wouldn't differentiate. Spemann thought he had support for this hypothesis when he did transplantation experiments in newts. When he transplanted the dorsal lip from one newt to another, he got conjoined twin newts - suggesting that some signal from the dorsal lip was indeed patterning the main body axis12,13.
Soon Spemann revised his hypothesis, largely based on careful observation of gastrulating embryos and on the cross-species transplantation work by Mangold. Mangold took the blastopore lip from an unpigmented newt species and transplanted under the ventral ectoderm of an early gastrula pigmented newt from the same genus. In this way she was able to see which tadpole tissues came from the donor (the unpigmented newt) and which from the host (the pigmented newt). They found that only a tiny portion of the twinned tadpole tissue was unpigmented - the notochord (plus some additional cells here and there - the technique was not completely perfect). This meant two important things: 1) The dorsal lip of the blastopore develops into the notochord. 2) The notochord patterns the tissue around it to make the primary body axis15.
Gastrulation in Evolution
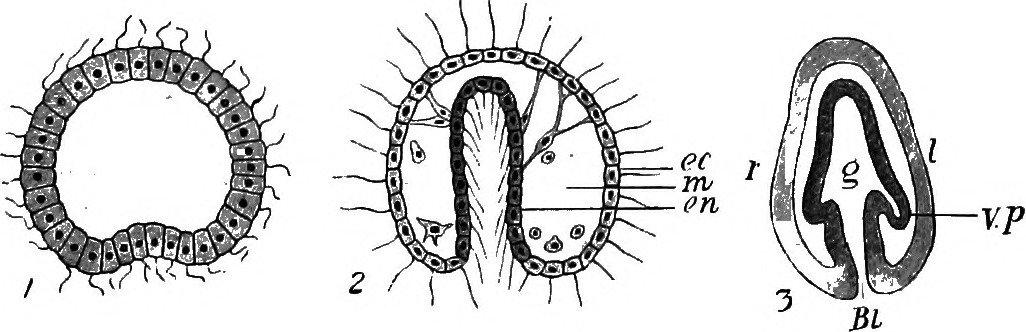
In general, gastrulation involves the involution and invagination of outer-layer cells to become internalized. Other outer layer cells epibolize to cover these internalizing cells. In this way, an embryo goes from a single cell type to multiple distinct cell layers. The place where this invagination or involution (where cells internalize) occur is called the blastopore. The blastopore can be a discrete "hole" in the embryo that elongates (called an archenteron) as in the case of sea urchins (Figure 7), or it can be a broad lip of cells that narrows as epibolizing cells expand as in frogs and snails (Figures 5 and 8).
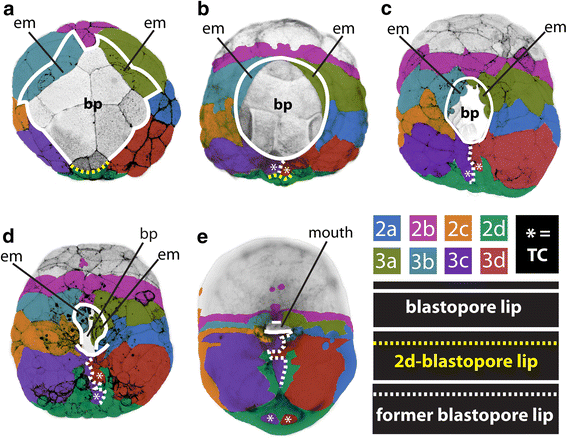
How can two such different structures be compared and even homologized? We consider gastrulation and blastopore formation to be homologous across Planulozoa (the group containing most animals, Figure 9) for two main reasons:
- Other multicellular organisms do not undergo the massive tissue rearrangements we see in animal development. In this way, gastrulation represents an animal (or at least a Planulozoa) synapomorphy.
- There are commonalities between various modes of gastrulation. The initial site of gastrulation is nearly always opposite the site of polar body extrusion, on the vegetal pole of the animal11. And of course the end result of gastrulation - a tube with ectoderm lining the outside and endoderm lining the inside with mesoderm or mesenchyme in the middle - is clearly homologous among planulozoans.