
9.2: Mutualism
- Page ID
- 25471

At the deepest level, the eukaryotic cell seems to have been created of interactions among separate prokaryotic species one or two billion years ago. Mitochondrial genomes are separate genomes within the cell with parallels to bacterial genomes, but mitochondria are no longer able to live on their own except under very special circumstances. Chloroplasts are similar. These are mutualisms at the very basis of complex life. Other mutualisms, like those with gut bacteria, form higher levels.
Mutualisms among pollinators and flowers are an ingenious arrangement for the fertilization of immobile organisms. A bee, for example, collects pollen as a food source and in the process spreads pollen from plant to plant. Plants advertise the pollen with bright flowers, and provide sweet nectar as an additional attraction (Figure \(\PageIndex{1}\), left).

A small mammal can act similarly, drinking nectar from one plant, getting pollen on its whiskers, and transferring the pollen when it drinks from another plant, in turn pollinating that plant (Figure \(\PageIndex{1}\), middle).
A fascinating pollination mutualism extends across the wetlands of the North American Upper Midwest, in marsh milkweed, Asclepias incarnata (Figure \(\PageIndex{1}\), right). Mutualisms are not necessarily perfect, and each member can be exploited in some small way. In this case, pollinators land on the milkweed flower and stand on “landing platforms” while taking nectar. But the platforms are not secure; they are curved and slightly difficult footholds. A pollinator’s legs slip into cracks between the platforms and, when the pollinator pulls its leg out, there are “saddle bags” of pollen (pollinia) wrapped around its leg, which can’t be dislodged. As it flies to a another flower, the saddle bags rotate forward so they easily dislodge, and they’re pulled off when the pollinator slips on another insecure landing platform of the corresponding species.
In another example, warthogs (Figure \(\PageIndex{2}\) left) attract abundant ticks. What better place, then, for Oxpeckers to find morsels of food than in the bristly fur of a water buffalo? A little blood in the tick is probably a bonus. This is good for the water buffalo, good for the oxpecker, and not good for the tick. This three-species interaction is (1) predation on the water buffalo by the tick, (2) predation on the tick by the oxpecker, and thus (3) mutualism between water buffalo and oxpecker. It is an “enemy of my enemy is my friend” interaction, one of the forty kinds of three-species interactions you will see in upcoming chapters.

Likewise, ants ward off potential predators from aphids (Figure \(\PageIndex{2}\) middle), and “cleaner fish” swim freely within the mouth of a large fish (Figure \(\PageIndex{2}\) right) while they remove the fish’s parasites.
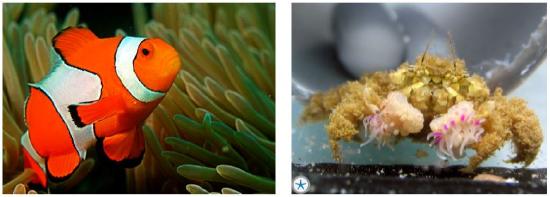
Sea anemones have stinging tentacles that other fish must avoid, but clown fish can resist the sting (Figure \(\PageIndex{3}\) left). This mutualism is more complex. The clown fish protect themselves from predators by living among the anemones, and their bright colors may attract predators who become prey for the anemone. Clown fish eat scraps of food missed by the anemone, plus their own food from the water column, and provide nitrogen to the anemone from their digested waste. These are intricate mutualisms, in which specifically matched species of clown fish and anemone have become permanent partners.
Pom-pom crabs employ sea anemones as weapons (Figure \(\PageIndex{3}\), right), carrying two anemones and waving them in a dance to dissuade approaching predators. Crabs are sloppy eaters, so the sea anemones get the benefit of the mess as payment.

Early on, our hunter–gatherer predecessors were predators, but later they domesticated some of their prey, changing some predator–prey interactions to mutualisms. From the point of view of domesticated sheep (Figure \(\PageIndex{4}\), left), humans may not be ideal mutualists. We protect them from wolves, harbor them from disease, and shield them from the worst vagaries of weather. But we also confine them to pens, shear off their wool, and kill and eat their lambs. Yet as agriculture advanced, the more people there were on the planet, and the more sheep there were, and vice versa. This is the ecological making of a mutualism.
It is similar with crops. Instead of gathering grain and fruit from forest and field, we cleared areas specifically for domesticated plants. We protect the crops from competition with other plants, work to keep them free of disease, and add water and nutrients to the soil to help them grow. For thousands of years we used beasts of burden to accelerate cultivation, also in mutualisms with those beasts. Now we are entering a wholly synthetic phase, in which teams of satellite-guided tractors navigate fields with the power of a thousand horses (Figure \(\PageIndex{4}\) right).
For a thought exercise, you may want to ponder our relationship with machines. If we were mutualists with draft horses, are we now mutualists with the machines that have replaced them? As you proceed through the next chapters, consider whether our relationships with our machines meet the ecological requirements of mutualisms.