
22.6: Architecture of Genome Organization
- Page ID
- 41055

Multiple cell types influence on determining architecture
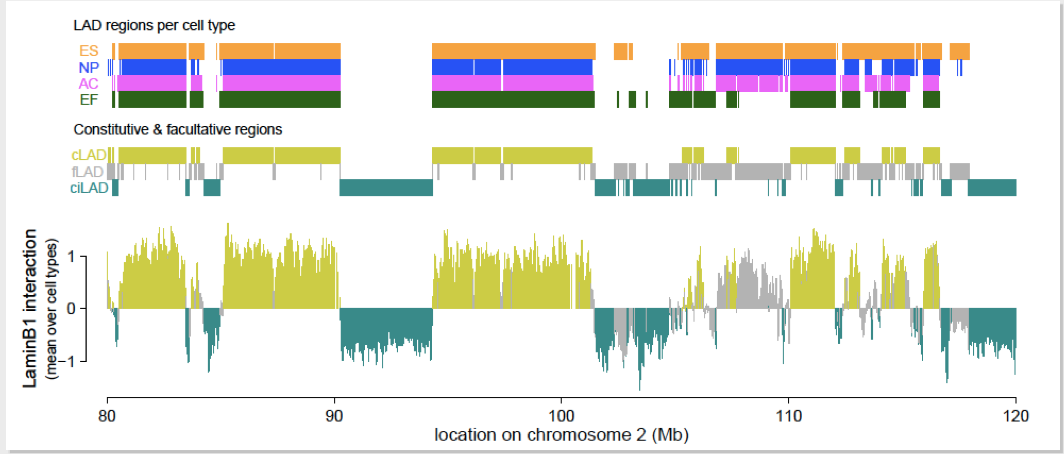
Embryonic stem cells (ESC), Neural Progenitor Cells (NPC), and Astrocytes(AC) are all isogenic cell types (they all start as embryonic stem cells). Embryonic stem cells are constantly dividing and are completely undifferentiated; they generate the neural progenitor cells, which are still dividing but less so, and are only halfway differentiated. The neural progenitor cells then generate the completely differentiated astrocytes. It was discovered that during this differentiation process, some areas switched from being Lamina Associated Domains to being interior domains. In the embryonic stem cells, there is very little transcription. However, transcription goes up as the cells become more and more differentiated. This matches the localization of the domains from being primarily associated with the lamina (and thus not expressed) to being localized to the interior. Even though these cell types each have very different properties, a DamID map shows a high level of similarity between the three isogenic cells as well as an independent fibroblast cell. Hidden Markov Models were employed to identify the Lamina Associated Domains between the cells. A core chromosome architecture was found with about 70% of the chromosome being constitutive (cLad/ ciLAD) and 30% of the chromosome being facultative(fLAD).
Inter-species comparison of lamina associations
To determine lamina associations between species, a mouse and a human were used. A genome wide alignment was constructed between a mouse and a human. For each genomic region in the mouse, the best reciprocal region was matched in the human. Then the human genome was re-mapped, and used to reconstruct a mouse genome. DamID data was projected onto this map and there was 83% concordance between the two genomes (91% for the constitutive regions; 67% for the facultative regions).
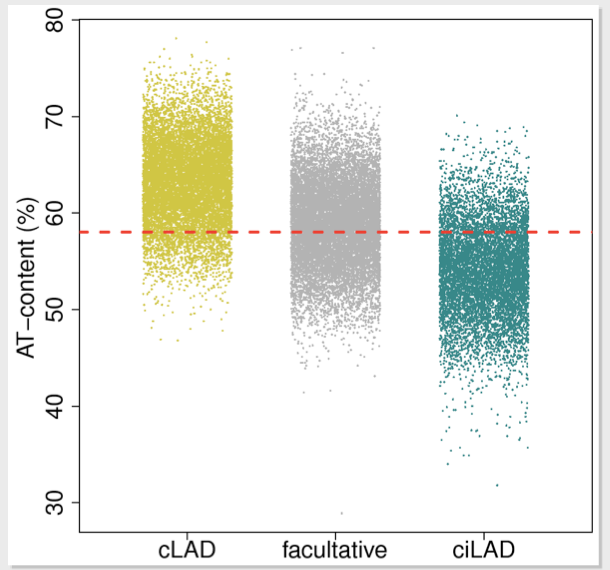
A-T Content Rule
A-T content has been found to be a strong predictor for lamina association within core architecture. Addi- tional support for this prediction is that the LAD-structure that makes up the core architecture is similar to an isochore structure (a large uniform region of DNA).
Cold Spring Harbor Laboratory Press. All rights reserved. This content is excluded from our Creative
Commons license. For more information, see http://ocw.mit.edu/help/faq-fair-use/.
Source: Meuleman, Wouter, et al. "Constitutive Nuclear Lamina-genome Interactions are Highly Conserved
and Associated with A/T-rich Sequence." Genome Reserach 23, no. 2 (2013): 270-80.