
6.4: Gluconeogenesis
- Page ID
- 16124

Having considered the initial anabolic reaction of life - carbon fixation by photosynthesis, we now turn our attention to utilizing the smaller metabolites to generate glucose and other sugars and carbohydrates. Glucose is the most important fuel for most organisms, and the only fuel for some cell types, such as brain neurons. Potential building blocks of glucose include many of the products and intermediates of glycolysis and the TCA cycle, as well as most amino acids. The key reaction is conversion of any of these compounds into oxaloactetate before using them to make glucose. In animals, the amino acids leucine and isoleucine, as well as any fatty acids, cannot be used to build glucose because they convert first to acetyl-CoA, and animals have no pathway for acetyl-CoA to oxaloacetate conversion. Plants, on the other hand, can push acetyl-CoA to oxaloacetate through the glyoxylate cycle, which will be discussed shortly.
The process of gluconeogenesis is in many ways the simple opposite of glycolysis, so it is not surprising that some of the enzymes used in glycolysis are the same as those used for gluconeogenesis. However, there are a few exceptions. These arose (and have probably evolved) for two major reasons -
- the thermodynamics of the reaction are prohibitive, and
- the need for independent control of the catabolic and anabolic processes.
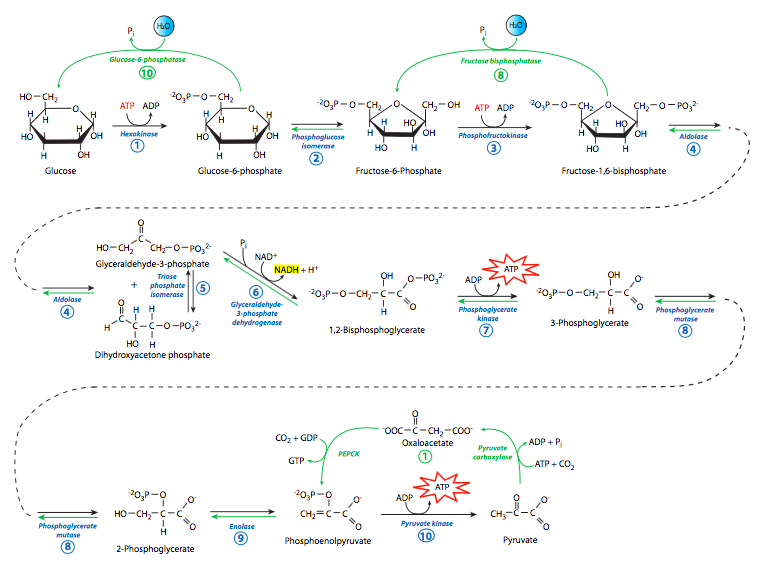
Since there is this parallel, we will explore gluconeogenesis first by starting with one of the major products of glycolysis, pyruvate. Pyruvate can be converted to oxaloacetate by pyruvate carboxylase, in a reaction requiring ATP hydrolysis. The oaxaloacetate is then converted to phosphoenolpyruvate (PEP) by PEP carboxykinase, which also uses nucleotide triphosphate hydrolysis for energy, though this time it is GTP.
Interestingly, PEP carboxykinase (PEPCK) is unregulated at the protein level. There are no known activators or inhibitors of its activity. The only regulation of PEPCK appears to be at the level of transcription: glucagon can stimulate it (as can gluocorticoids and thyroid hormone), while insulin can inhibit it. The other gluconeogenic enzymes, though, do have direct activators and inhibitors. They are allosteric modulators, binding away from, but influencing the shape and efficacy of the substrate binding site. In examining the regulation of these enzymes, one important regulator stands out because it is not a metabolite of either glycolysis or gluconeogenesis. Fructose-2,6-bisphosphate (F2,6P) is an activator of phosphofructokinase, and an inhibitor of fructose bis-phosphatase. F2,6P levels are controlled by fructose-bis-phosphatase-2 and phosphofructokinase-2, which are themselves controlled by levels of fructose-6-phosphate, as well as through a hormone-driven signaling cascade shown in the Figure on the next page.
As shown in the summary/comparison (Figure \(\PageIndex{8}\)), from the formation of PEP to the formation of fructose-1,6-bisphosphate the enzymes used in gluconeogenesis are exactly the same enzymes used in glycolysis. This works because the free energy change in those reactions is relatively small. However, in the dephosphorylation of fructose-1,6-bisphosphate to to fructose-6-phosphate, and subsequently in the dephosphorylation of glucose-6-phosphate to glucose, there is a large free energy change that works against the gluconeogenic reactions. Thus, the enzymes that drive these reactions are different from the enzymes that drive the reverse reactions in glycolysis (i.e. hexokinase, phosphofructokinase). These two hydrolytic reactions are catalyzed by fructose bis-phosphatase and glucose-6-phosphatase, respectively. Full reversal of glycolysis in animals is limited, however, to liver and kidney, since they are the only tissues that express glucose-6-phosphatase. Other tissues use different mechanisms for generating glucose (e.g. glycogenolysis).
The glyoxylate cycle provides a mechanism for plants to convert acetyl-CoA into oxaloacetate, and therefore contribute to gluconeogenesis. This allows them to convert fatty acids and the hydrophobic amino acids leucine and isoleucine into glucose when necessary. The ability to do this comes from a plant-specific organelle called the glyoxysome, as well as some mitochondrial enzymes. The glyoxysomal part of the cycle consists of ve steps, of which the first three contribute to the conversion, while the last two steps regenerate the glyoxysomal oxaloacetate (Figure \(\PageIndex{9}\)).
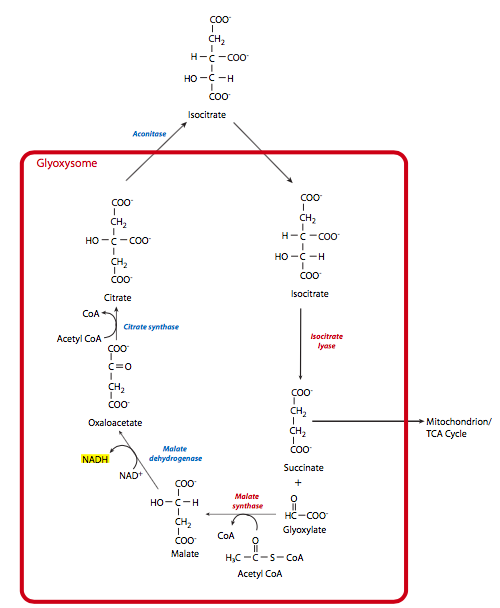
- Once the macromolecules have been broken down to acetyl-CoA, they enter the glyoxysome and combine with oxaloacetate to make citrate. This is catalyzed by citrate synthase just as in the mitochondrial TCA cycle. The next reaction also uses a familiar enzyme: aconitase catalyzes the conversion of citrate to isocitrate. However, the aconitase is a cytosolic enzyme, so the citrate is transported out of the glyoxysome and then the isocitrate transported back in.
- At this point, the glyoxysomal-specific enzyme, isocitrate lyase, hydrolyzes isocitrate to yield succinate and glyoxylate. The succinate is transported to the mitochondrion, where TCA cycle enzymes convert it to fumarate and then malate, which is transported out to the cytosol. In the cytosol, the malate is converted to oxaloactetate through malate dehydrogenase, and gluconeogenesis can proceed.
- The glyoxylate is acted upon by another glyoxysomal enzyme, malate synthase, which adds it to acetyl-CoA to form malate.
- The final step of the glyoxysomal portion of the glyoxylate cycle is oxidation of the malate to oxaloacetate by glyoxysomal malate dehydrogenase.
So, to summarize, the pool of oxaloacetate within the glyoxysome is used and regenerated within the glyoxysome. Acetyl-CoA is converted to succinate within the glyoxysome, but then goes to the mitochondrion for conversion to malate, and finally the cytosol for conversion to a separate pool of oxaloacetate that is then used in gluconeogenesis.