
1: Anatomy of a Cell - A Very Brief Overview
- Page ID
- 16086

Since this entire course is devoted to understanding the workings of the cell, it is almost superfluous to dedicate a chapter to identifying the parts of the cell and their functions. However, because it is easy to get lost in the intricacies of the molecules and chemical reactions within the cell, consider this chapter more of a framework or map for the course, giving context to the minutiae.
The cell is the smallest unit of life, so all cells, whether they are unicellular organisms or just a tiny part of a multicellular organism, have certain characteristics in common: they must contain genetic information and the mechanisms to regulate and use that information to produce its own parts and to reproduce new cells, they must be able to use energy in chemical reactions and physical actions, they must be able to regulate those activities, and they must respond to stimuli.
Cells use DNA (deoxyribonucleic acid) for their genetic material, and all cells contain the transcriptional and translational enzymes to read it and use the information to construct more cell components. However, simply having genetic material does not define life: viruses have genetic material containing all the information necessary to make a complete virus, but it does not contain the enzymes necessary to do so, nor the ability to obtain the raw molecular material needed to do so. It is absolutely dependent on the machinery inside whatever cell it infects. Therefore, a virus is not a living organism.
The genome is not only an instruction set for making a cell (or an organism, for that matter); it is also replicable itself. Roughly speaking, during part of its life cycle, the cell makes an extra copy of its genome and increases the numbers of all the other “stuff” (proteins, fats, etc.) of which it is made, and then it reproduces by division. The mother cell splits into two daughter cells, each with the same complement of genetic information, and with approximately the same cellular components. Thus we see that while the genome is often considered the blueprint for a cell/organism, in fact cells are not built up from scratch directly from DNA. Every cell comes from another cell. The DNA can then be used to customize that cell for specific purposes as determined by its environment. When a particular component of the cell is needed, the information for making that component is read from the DNA and copied into RNA which is used as a program from which ribosomes can manufacture the proteins needed. A living cell needs all these things: the genetic information, the mechanisms and machinery to use the information to build cell parts, and the ability to harness energy to do so.
As we will see in chapter 3, the physical laws of nature require that everything tends towards its simplest, least organized, state unless there is an input of energy to work against that tendency. Since cells are a highly ordered collection of very complex molecules, they must therefore require energy to remain as cells. Thus, life requires the ability to obtain energy, either from sunlight or food, and the ability to convert that energy into forms that can be readily used by the cell to maintain itself by building or rearranging necessary molecules and macromolecular structures.
How do cells know when to carry out these activities? This leads us to the next characteristic of living cells: the ability to respond to stimuli. In other words, they are self-regulating. If glucose levels run low and the cell needs energy, glucose transport proteins are made, or if the cell needs to move to an area of higher food concentration, the cell cytoskeleton rearranges to move the cell. The cell has the ability to initiate repair processes if it detects lesions in its genome, it can pause the cell cycle to allow such repair processes time, and it can even initiate its own death if repairs are repeatedly unsuccessful.
In addition to responding to internal signals, living cells are also able to respond to external stimuli. Whether it is contact with a neighboring cell, binding a hormone released from a cell far away, or simply interacting with non-cellular environmental objects, a cell is able to respond to such stimuli. Responses may include making new proteins, destruction of existing proteins, moving away from the stimulus, moving towards the stimulus, initiation of reproduction, and many other possibilities.
There are two basic types of cells: prokaryotes and eukaryotes. The difference is simple and readily recognizable under light microscopy. Eukaryotic cells contain intracellular membrane-bound compartments (called organelles). Prokaryotic cells do not contain any such compartments (Figure \(\PageIndex{1}\)).
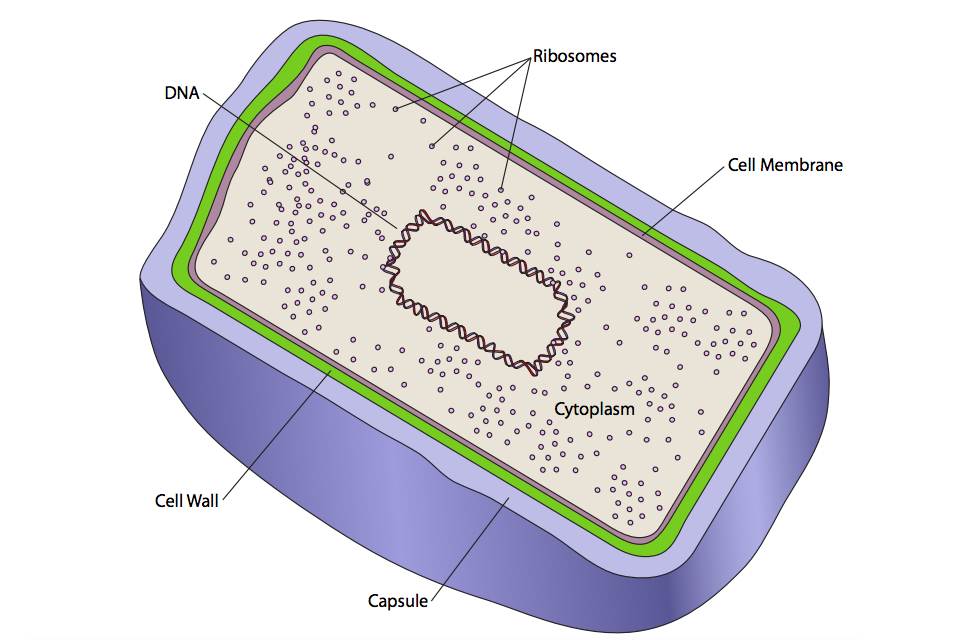
There is only one membrane in prokaryotes, the cell membrane, and only one compartment in prokaryotic cells, the cytoplasm. That does not preclude a certain level of organization in prokaryotes, but it is not as complex as eukaryotes. The genomic DNA is usually organized in a central nucleoid. There are not intracellular membranous organelles, but the cell is defined by a cell membrane. Outside of the cell membrane, prokaryotes have a cell wall. This wall is relatively rigid and confers shape to the cell. Depending on the type of bacteria, the thickness of the wall varies (thick = “gram positive”, thin = “gram negative”). Some, but not all bacteria also secrete another layer outside of the cell wall. This is a relatively tight matrix called a capsule that helps protect the cell from dessication in dry environments. A comparatively loose matrix of the same types of molecules may be secreted, and instead of the capsule, the result is called a slime layer. The slime layer is important in bacterial attachment and formation of biofilms (see chapter 13, Extracellular Matrix).
Eukaryotic cells are considerably more complex. Eukaryotic organisms are currently classified into four kingdoms: animal, plant, fungus, and protists. The animal cell in Figure \(\PageIndex{2}\) depicts has many features in common with cells of the other three kingdoms.
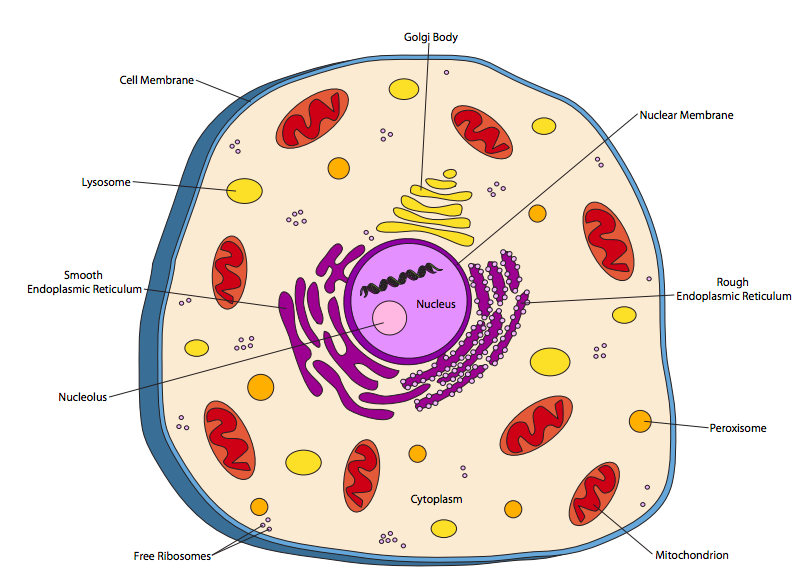
Obviously, the biggest difference between the animal cell (or any eukaryotic cell) and prokaryotic cells is the presence of internal membrane-bound compartments, or organelles. The most prominent of these is the nucleus, which houses the DNA. Traditionally, it has been assumed that most eukaryotic genomes can range from 10 to 100 x 106 nucleotides (10-100 Mb) in total length, over two or more chromosomes (DNA molecules) of roughly similar size. In contrast, prokaryotic genomes have traditionally been viewed as a single circular chromosome, and mostly under a megabase (106 nucleotides) in length.
Recent and better methods for genome mapping and sequencing, and a broadening of the sample organisms has shown those numbers to be inaccurate. In fact, eukaryotic genomes range from ~3 Mb to over 4000 Mb. Prokaryotic genomes vary from 0.5 Mb to a little over 10 Mb (0.5 to 6 Mb for Archaea, 0.6 to 10 Mb for Bacteria) and may be spread over multiple DNA molecules that may be either linear or circular.
The nucleus is bounded by a double-layered membrane (most other organelles are bounded by a single membrane) that is continuous with the the endoplasmic reticulum (ER). The endoplasmic reticulum is subdivided into the rough ER (RER) and the smooth ER (SER) based on appearance in electron micrographs. The “studs” on the RER are ribosomes, which are the molecular machinery for making proteins in the cell. There are also free-floating ribosomes - the difference is that the free ribosomes make proteins that stay in the cytoplasm, while ribosomes attached to the RER are synthesizing proteins that are destined to insert into a membrane, localize inside an organelle, or be secreted out of the cell entirely. The RER makes modifications to the proteins as well as compartmentalizing them. The SER counts lipid synthesis (e.g. to make membranes) and detoxification reactions among its duties. It should be noted that ribosomes on the RER are not permanently attached, and after they have produced a protein, they dissociate from the RER and rejoin the general pool of free ribosomes in the cytoplasm.
The Golgi complex, or Golgi bodies, while physically independent, are a functional extension to the protein processing and sorting that occurs in the ER. Proteins leave the Golgi in vesicles bound for the cell membrane or other organelles. Vesicles, while membrane-bound, are not generally counted as organelles: they are simply small transport packages.
Mitochondria are complex organelles that are not only bounded by a membrane, but also contain a second membrane that is highly crenulated. Mitochondria make aerobic respiration possible, using oxygen as an oxidizer to produce chemical energy (i.e. ATP) far more efficiently than the anaerobic processes used by most prokaryotes. This ability to produce more energy from the same amount of food allows eukaryotic cells to grow larger than prokaryotes.
Lysosomes are acidic and contain digestive enzymes that break down large food molecules — particularly proteins and fats — to make them usable by the rest of the cell. These enzymes work optimally in acidic conditions, which acts as a sort of safety mechanism: if a lysosome breaks and releases its enzymes into the cytoplasm, they will not break down cellular components willy-nilly because the cytoplasmic pH is close to neutral and the enzymes do not work well. Once thought to be exclusive to animal cells, lysosomes have now been described in all cells from all eukaryotic kingdoms.
Peroxisomes also break down or convert molecules, but they generally act on smaller molecules by oxidation. For example, some peroxisomes in human liver cells are used to break down alcohol (ethanol). Processes like this often produce H2O2, hydrogen peroxide, as a byproduct. Since H2O2 in high concentrations is harmful, peroxisomes often contain an enzyme, catalase, that converts it into water and molecular oxygen.
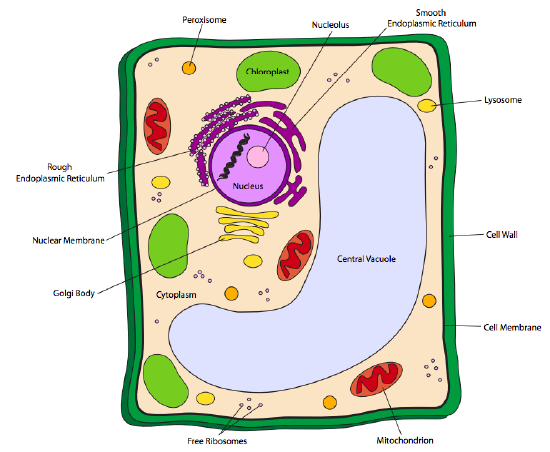
Plant cells have all of the above named organelles, but additionally may also bear two other types of organelles: chloroplasts and vacuoles. In addition to this, plant cells also have a rigid cell wall external to the cell membrane. Chloroplasts are similar to mitochondria in shape and structure (membranes within the outer membrane). However, its function is very different: chloroplasts absorb light energy from the sun (or other light sources) and convert them into chemical energy in the form of simple sugars for the cell to store and use later, in essence turning the cell into a solar energy warehouse and distribution center. Since plants and some photosynthetic bacteria are the only organisms capable of converting solar energy into a form useful to living cells, they are crucial to the survival of all other life.
Vacuoles are essentially storage units. They may store starches for use as energy sources when sunlight is unavailable or when immediate photosynthesis alone is not sufficient to provide for the energy needs of the cell. Other vacuoles, such as the one depicted above in Figure \(\PageIndex{3}\), store water, which helps the cell to maintain rigidity in combination with the cell wall.
Plant cell walls are composed of very different materials than the previously mentioned bacterial cell walls. Plant cell walls are primarily composed of the glucose polymer, cellulose, but contain other polysaccharides as well. Depending on the type of plant cell, there may be multiple layers of cellulose composing the cell wall. The wood and bark of trees, for example have both a primary (thin) cell wall and a secondary (thick) wall, while the leaves would have only a primary wall. Fungi also have cell walls, and they too are different from bacterial cell walls. True fungi have cell walls that are composed primarily of the polysaccharide chitin, and no cellulose.
Interestingly, two groups once classified as fungi: oomycetes and dictyostelids, have cell walls composed of cellulose (and some have both). These organisms have been reclassified in Protista.
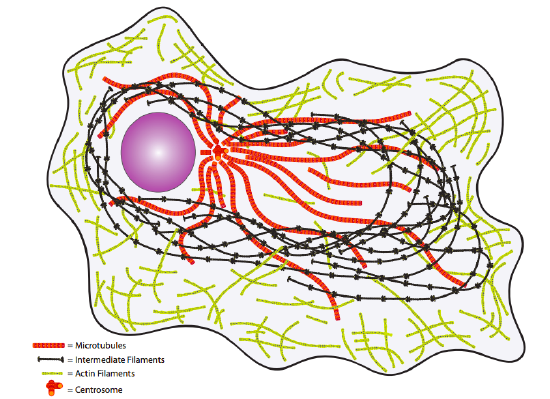
Finally, consider the cytoplasm. Once considered merely the aqueous environment in which the “important” molecules or organelles floated, it is now better understood to be filled with important structural and transport elements (Figure \(\PageIndex{4}\)). The cytoskeleton provides not only an internal physical structure but also a transport system to move molecules, vesicles, and even organelles to where they are needed.
All of the cell parts introduced in this chapter will be explained in much greater detail in subsequent chapters. More importantly, the intertwined relationships between many of the molecules and organelles will be discussed and elucidated. As you go through this course, you will notice that the same species come up over and over as examples. These are the model organisms upon which the great majority of molecular cell biology research is based. Most prokaryotic research has been based on Escherichia coli (E. coli), which is a Gram-negative rod-shaped bacterium commonly found in the gut of many higher animals. The Gram-negative soil bacteria, Bacillus subtilis, is a sporeforming organism that has also been used in research because, like E. coli, its genome is easily manipulated for experimentation, and is also relatively easy to grow in the lab. On the eukaryotic side, yeast (Saccharomyces cerevisiae, or Schizosaccharomyces pombe) are very commonly used for simpler intracellular processes due to simple genetics and very fast generation times. Caenorhabditis elegans (a nematode) and Drosophila melanogaster (fruit fly) are popular invertebrate model organisms, especially for developmental and genetic studies due to the small number of cells, mostly with traceable lineage, and fast generation time (for metazoans). Frogs, particularly the South African clawed frog, Xenopus laevis, and the Northern Leopard frog, Rana pipiens, are popular for certain types of developmental and cell cycle studies because they have huge oocytes that are amenable to many kinds of genetic and physiological manipulation not possible in other cells. Arabidopsis thaliana is the most commonly used model organism for the study of plant genetics. Finally, because they are mammals like us (humans), but breed quickly and can be genetically manipulated with relative ease, mice (Mus musculus) are very commonly used in the study of more complex intra- or inter-cellular mechanisms. More recently, the near complete sequencing of the genome and development of techniques to manipulate it, have made the rat (Rattus norvegicus) another viable research organism for the study of mammalian genes.
Further information on model organisms can be found at the United States’ National Institutes of Health web site: www.nih.gov/science/models/
The commonalities that make all of these organisms excellent models for the study of the molecules of the cell and the interactions between them that constitute life, are a relatively short generation time, well-described (and in most cases fully sequenced) genome, and ease of experimental manipulation. Most of the molecules and mechanisms you will learn in the course were discovered in the simpler model mechanisms, and then found again, often with elaboration in the more complex ones.

Thumbnail: A diagram of a typical prokaryotic cell. (Public Domain; LadyofHats).