
4.3: Membrane Transport Proteins
- Page ID
- 16109

Membrane proteins come in two basic types: integral membrane proteins (sometimes called intrinsic), which are directly inserted within the phospholipid bilayer, and peripheral membrane proteins (sometimes called extrinsic), which are located very close or even in contact with one face of the membrane, but do not extend into the hydrophobic core of the bilayer. Integral membrane proteins may extend completely through the membrane contacting both the extracellular environment and the cytoplasm, or they may only insert partially into the membrane (on either side) and contact only the cytoplasm or extracellular environment. There are no known proteins that are completely buried within the membrane core.
Integral membrane proteins (Figure \(\PageIndex{9}\)) are held tightly in place by hydrophobic forces, and purification of them from the lipids requires membrane-disrupting agents such as organic solvents (e.g. methanol) or detergents (e.g. SDS, Triton X-100). Due to the nature of the bilayer, the portion of integral membrane proteins that lie within the hydrophobic core of the membrane are usually very hydrophobic in character, or have outward-facing hydrophobic residues to interact with the membrane core. These transmembrane domains usually take one of the two forms depicted in Figures 8 and 14: alpha helices - either individually or in a set with other alpha helices, or barrel-shaped insertions in which the barrel walls are constructed of beta-pleated sheets. The hydrophobic insertions are bounded by a short series of polar or charged residues that interact with the aqueous environment and polar head groups to prevent the hydrophobic portion of the protein from sliding out of place. Furthermore, proteins can have multiple membrane- spanning domains.
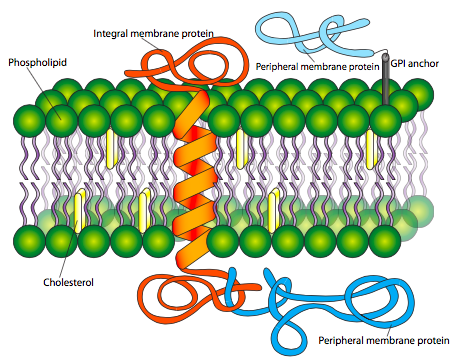
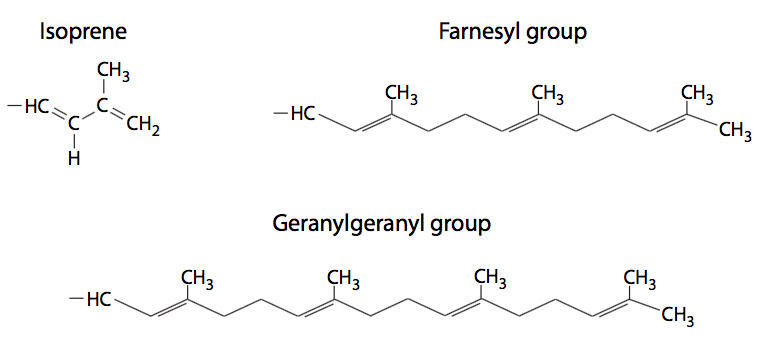
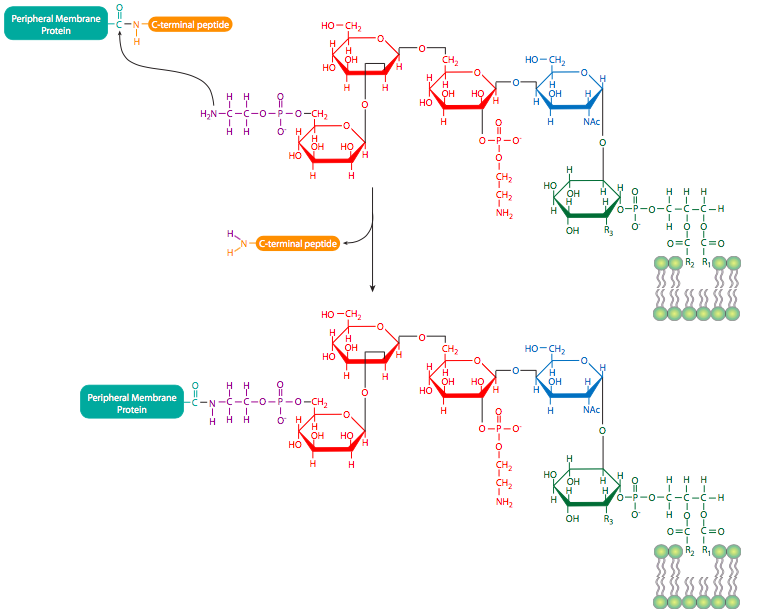
Peripheral membrane proteins (also shown in Figure \(\PageIndex{9}\)) are less predictable in their structure, but may be attached to the membrane either by interaction with integral membrane proteins or by covalently attached lipids. The most common such modifications to peripheral membrane proteins are fatty acylation, prenylation, and linkage to glycosylphosphatidylinositol (GPI) anchors. Fatty acylation is most often a myristoylation (a 14:0 acyl chain) and palmitoylation (a 16:0 chain) of the protein. A protein may be acylated with more than one chain, although one or two acyl groups is most common. These fatty acyl chains stably insert into the core of the phospholipid bilayer. While myristoylated proteins are found in a variety of compartments, almost all palmitoylated proteins are located on the cytoplasmic face of the plasma membrane. Prenylated proteins, on the other hand, are primarily found attached to intracellular membranes. Prenylation is the covalent attachment of isoprenoids to the protein - most commonly isoprene (a C5 hydrocarbon), farnesyl (C15), or geranylgeranyl (C20) groups (Figure \(\PageIndex{10}\)). GPI anchors (Figure \(\PageIndex{11}\)) are found exclusively on proteins on the outer surface of the cell, but there does not appear to be any other commonality in their structures or functions.
Of course, not all membrane proteins, or even all transmembrane proteins, are transporters, and the many other functions of membrane proteins - as receptors, adhesion molecules, signaling molecules, and structural molecules - will be discussed in subsequent chapers. The focus here is on the role of membrane proteins in facilitating transport of molecules across the cell membrane.
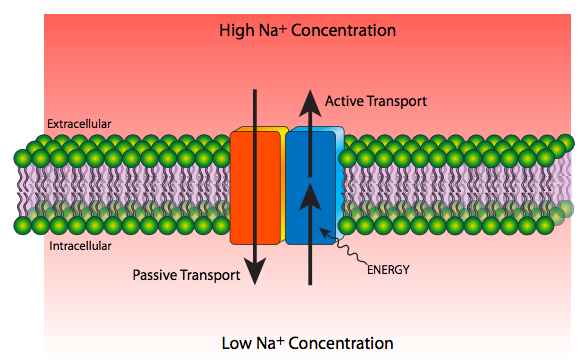
Transport across the membrane may be either passive, requiring no external source of energy as solute travels from high to low concentration, or active, requiring energy expenditure as solute travels from low to high concentration (Figure \(\PageIndex{12}\)).
Passive transport can also be divided into nonmediated transport, in which the movement of solutes is determined solely by diffusion, and the solute does not require a transport protein, and mediated passive transport (aka facilitated diffusion) in which a transport protein is required to help a solute go from high to low concentration. Even though this may sometimes involve a change in conformation, no external energy is required for this process. Nonmediated passive transport applies only to membrane-soluble small nonpolar molecules, and the kinetics of the movement is ruled by diffusion, thickness of the membrane, and the electro-chemical membrane potential. Active transport is always a mediated transport process.
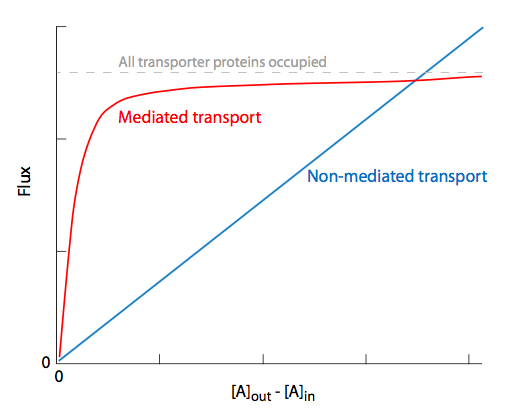
Comparing the solute flux vs initial concentration in Figure \(\PageIndex{13}\), we see that there is a linear relationship for nonmediated transport, while mediated passive transport (and for that matter, active transport) shows a saturation effect due to the limiting factor of the number of available proteins to allow the solute through. Once there is enough solute to constantly occupy all transporters or channels, maximal ux will be reached, and increases in concentration cannot overcome this limit. This holds true regardless of the type of transporter protein involved, even though some are more intimately involved in the transport than others.
In addition to protein transporters, there are other ways to facilitate the movement of ions through membranes. Ionophores are small organic molecules, often (but not exclusively) made by bacteria, that help ions move through membranes. Many ionophores are antibiotics that act by causing the membranes to become leaky to particular ions, altering the electrochemical potential of the membrane and the chemical composition inside the cell. Ionophores are exclusively passive-transport mechanism, and fall into two types.
The first type of ionophore is a small mostly-hydrophobic carrier almost completely embedded in the membrane, that binds to and envelopes a speci c ion, shielding it from the lipid, and then moves it through the cell membrane. The most studied carrier-type ionophore is valinomycin, which binds to K+. Valinomycin is a 12-residue cyclic depsipeptide (contains amide and ester bonds) with alternating d- and l- amino acids. The carbonyl groups all face inward to interact with the ion, while the hydrophobic side chains face outward to the lipid of the membrane. Carrier ionophores are not necessarily peptides: the industrial chemical 2,4-dinitrophenol is an H+ carrier and important environmental waste concern, and nystatin, an antifungal used to treat Candida albicans infections in humans, is a K+ carrier.
The second type of carrier forms channels in the target membrane, but again, is not a protein. Gramicidin is a prototypical example, an anti-gram-positive antibacterial (except for the source of gramicidins, the gram-positive Bacillus brevis) and ionophore channel for monovalent cations such as Na+, K+, and H+. It is im- permeable to anions, and can be blocked by the divalent cation Ca2+. Like valinomycin, gramicidin A is also a made of alternating d- and l- amino acids, all of which are hydrophobic (l-Val/ Ile-Gly-l-Ala-d-Leu-l-Ala-d-Val-l-Val-d-Val-l-Trp-d-Leu-l-Trp-d-Leu- l-Trp-d-Leu-l-Trp). Gramicidin A dimerizes in the membrane to form a compressed b-sheet structure known as a b-helix. The dimerization forms N-terminal to N-terminal, placing the Trp res- idues towards the outer edges of the membrane, with the polar NH groups towards the extracellular and cytoplasmic surfaces, anchoring the pore in place.
Channels are essentially hands-off transport systems that, as the name implies, provides a passage from one side of the cell to another. Though channels may be gated - able to open and close in response to changes in membrane potential or ligand binding, for example - they allow solutes through at a high rate without tightly binding them and without changes in conformation. The solute can only move through channels from high to low concentration. The potassium channel depicted below (Figure \(\PageIndex{14}\)A) is an example: there is a selectivity lter (14B) of aligned carbonyl oxygens that transiently positions the K+ ions for rapid passage through the channel, but it does not bind the K+ for any significant period, nor does the channel undergo any conformational changes as a result of the interaction. Smaller Na+ ions could (and on rare occasion do) make it through the K+ channel, but because they are too small to be properly positioned by the K+ filter, they usually pop back out. It should be noted that this channel is a tetramer (14C) and the cutaway diagram in (14A) only shows half of the channel for clarity.
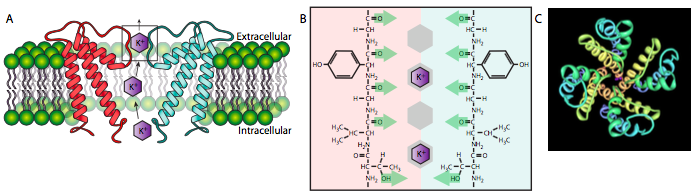
While most proteins called “channels” are formed by multiple alpha-helices, the porins are formed by a cylindrical beta sheet. In both cases, solutes can only move down the concentration gradient from high to low, and in both cases, the solutes do not make signi cant contact with the pore or channel. The interior of the pore is usually hydrophilic due to alternating hydrophilic/hydrophobic residues along the beta ribbon, which places the hydrophobic side chains on the outside, interacting with the membrane core.
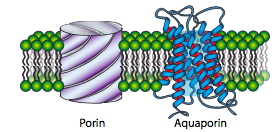
Porins are primarily found in gram-negative bacteria, some gram-positive bacteria, and in the mitochondria and chloroplasts of eukaryotes. They are not generally found in the plasma membrane of eukaryotes. Also, despite the similarity in name, they are structurally unrelated to aquaporins, which are channels that facilitate the diffusion of water in and out of cells.
Transport proteins work very differently from channels or pores. Instead of allowing a relatively fast ow of solutes through the membrane, transport proteins move solutes across the membrane in discrete quanta by binding to the solute on one side of the membrane, changing conformation so as to bring the solute to the other side of the membrane, and then releasing the solute. These transport proteins may work with individual solute molecules like the glucose transporters, or they may move multiple solutes. The glucose transporters are passive transport proteins, so they only move glucose from higher to lower concentrations, and do not require an external energy source. The four isoforms are very similar structurally but differ in their tissue distribution within the animal: for example, GLUT2 is found primarily in pancreatic b cells, while GLUT4 is found mostly in muscle and fat cells.
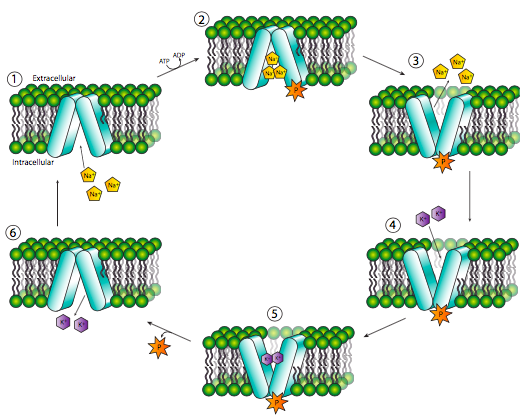
On the other hand, the classic example of an active transport protein, the Na+/K+ ATPase, also known as the Na+/K+ antiport, utilizes the energy from ATP hydrolysis to power the conformational changes needed to move both Na+ and K+ ions against the gradient. Referring to the Figure \(\PageIndex{16}\), in its resting state, the Na+/K+ ATPase is open to the cytoplasm and can bind three Na+ ions (1). Once the three Na+ have bound, the transporter can catalyze the hydrolysis of an ATP molecule, removing a phosphate group and transferring it onto the ATPase itself (2). This triggers a conformational change that opens the protein to the extracellular space and also changes the ion binding site so that Na+ no longer binds with high affinity and drops off (3). However, the ion binding site specificity is also altered in this conformational change, and these new sites have a high affinity for K+ ions (4). Once the two K+ bind, the attached phosphate group is released (5) and another conformational shift puts the transporter protein back into its original conformation, altering the K+ binding sites to allow release of the K+ into the cytoplasm (6), and revealing Na+ affinity once again.
The Na+/K+ ATPase is a member of the P-type family of ATPases. They are named because of the autophosphorylation that occurs when ATP is hydrolyzed to drive the transport. Other prominent members of this family of ATPases are the Ca2+-ATPase that pumps Ca2+ out of the cytoplasm into organelles or out of the cell, and the H+/K+ ATPase, though there are also P-type H+ pumps in fungal and plant plasma membranes, and in bacteria.
Cardiac glycosides (also cardiac steroids) inhibit the Na+/K+ ATPase by binding to the extracellular side of the enzyme. These drugs, including digitalis (extracted from the purple foxglove plant) and ouabain (extracted from ouabio tree) are commonly prescribed cardiac medications that increase the intensity of heart contractions. The inhibition of Na+/K+ ATPase causes a rise in [Na+]in which then activates cardiac Na+/Ca2+ antiports, pumping excess sodium out and Ca2+ in. The increased [Ca2+]cytoplasm is taken up by the sarcoplasmic reticulum, leading to extra Ca2+ when it is released to trigger muscle contraction, causing stronger contractions.
Unlike Na+ or K+, the Ca2+ gradient is not very important with respect to the electro- chemical membrane potential or the use of its energy. However, tight regulation of Ca2+ is important in a different way: it is used as an intracellular signal. To optimize the effectiveness of Ca2+ as a signal, its cytoplasmic levels are kept extremely low, with Ca2+ pumps pushing the ion into the ER (SR in muscles), Golgi, and out of the cell. These pumps are themselves regulated by Ca2+ levels through the protein calmodulin. At low Ca2+ levels, the pump is inactive, and an inhibitory domain of the pump itself prevents its activity. However, as Ca2+ levels rise, the ions bind to calmodulin, and the Ca2+-calmodulin complex can bind to the inhibitory region of the Ca2+ pump, relieving the inhibition and allowing the excess Ca2+ to be pumped out of the cytoplasm.
There are three other families of ATPases: the F-type ATPases are proton pumps in bacteria and mitochondria and chloroplasts that can also function to form ATP by running “backwards” with protons owing through them down the concentration gradient. They will be discussed in the next chapter (Metabolism). Also, there are V-type ATPases that regulate pH in acidic vesicles and plant vacuoles, and finally, there are anion-transporting ATPases.
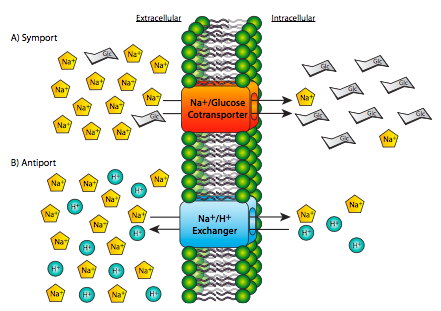
Hydrolysis of ATP, while a common source of energy for many biological processes, is not the only source of energy for transport. The active transport of one solute against its gradient can be coupled with the energy from passive transport of another solute down its gradient. Two examples are shown in Figure \(\PageIndex{17}\): even though one is a symport (both solutes crossing the membrane in the same physical direction) and one is an antiport (the two solutes cross the membrane in opposite physical directions), they both have one solute traveling down its gradient, and one solute traveling up against its concentration gradient. As it happens, we have used Na+ movement as the driving force behind both of these examples. In fact, the Na+ gradient across the membrane is an extremely important source of energy for most animal cells. However this is not universal for all cells, or even all eukaryotic cells. In most plant cells and unicellular organisms, the H+ (proton) gradient plays the role that Na+ does in animals.
Acetylcholine receptors (AchR), which are found in some neurons and on the muscle cells at neuromuscular junctions, are ligand-gated ion channels. When the neurotransmitter (acetylcholine) or an agonist such as nicotine (for nicotinic type receptors) or muscarine (for muscarinic type receptors) binds to the receptor, it opens a channel that allows the ow of small cations, primarily Na+ and K+, in opposite directions, of course. The Na+ rush is much stronger and leads to the initial depolarization of the membrane that either initiates an action potential in a neuron, or in muscle, initiates contraction.