
14.6: Roles of Transposition in Evolution and Diversity
- Page ID
- 16504

A. Transposons and Exon Shuffling
A role for unequal recombination in moving exons in and out of different eukaryotic split genes was described earlier. This kind of exon shuffling could happen when short DNA sequences in two different introns misalign during meiotic synapsis, allowing for unequal crossing over. Expression of a gene with a ‘new’ exon produces a protein with a new domain and a new activity. If the event is not harmful, diversity is increased!
When found in introns, transposons are long regions of DNA similarity that can stabilize synapsis, increasing the chances of unequal recombination and exon shuffling. For example, Alu (SINE) elements are often found within introns, where they can integrate with no ill effect. The similarity of Alu elements in the introns of unrelated genes does seem to account for exon shuffling by unequal crossing over between the different genes that share domains and specific functions as a result. Another way in which transposons facilitate germ line cell exon shuffling is more direct. Imagine a pair of transposons in introns of a gene on either side of an exon. Should such transposons behave like the two outer IS elements in a bacterial Tn element (discussed above), they might be excised as a single, large transposon containing an exon. The paired transposons flanking the exon might then insert in an intron of a completely different gene! This possibility is illustrated on the next page.
Transposon-mediated exon shuffling can explain insertion of exon-encoded domains of epidermal growth factor (EGF) into several unrelated genes. The mitogen EGF was discovered because it stimulated skin cells to start dividing. The gene for TPA (tissue plasminogen activator, a blood-clot dissolving protease) shares EGF gene domains. TPA is a treatment for heart attack victims that, if administered rapidly after the attack, can dissolve the clot and allow coronary artery blood flow to heart muscle to resume. Other genes that contain EGF domains include those for Neu and Notch proteins, both involved in cellular differentiation and development.
Some exon shuffling events may have been mediated by LINE transposition and by a special group of recently discovered transposons called helitrons. Helitrons replicate by a rolling circle mechanism. If you are curious about helitrons, do a google search to learn more about them, and what role they may have had in refashioning and reconstructing genomes in evolution. The general pathway of exon shuffling involving paired proximal DNA transposons is illustrated below.
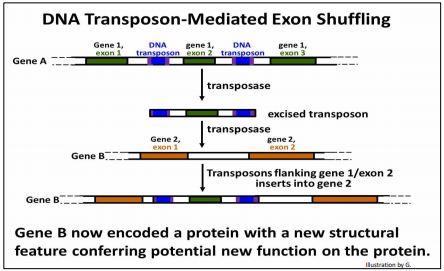
In the generic example shown above, exon 2 of gene A has been inserted, along with flanking transposons, into another gene (gene B).
B. Transposon Genes and Immune System and Genes Have History
Several important eukaryotic genes may have been derived from transposons. Perhaps the most intriguing example of this is to be found in the complex vertebrate immune system. Our immune system includes immunoglobulins (antibodies). You inherited genes for immunoglobulin proteins from your parents.
These genes contain multiple variant V, D, and J regions linked to a C region. V, D, J and C are defined as Variable, Joining, Diversity and Constant DNA regions, respectively. They will recombine to create many diverse V-D-J-C immunoglobulin antibody molecules (the D region is not always included in the final recombined gene). These gene rearrangements occur during the maturation of certain stem cells in bone marrow that will become immune cells (B or T lymphocytes). In response to a challenge by foreign substances called antigens, cells will be selected that contain rearranged immunoglobulin genes coding for immunoglobulins that can recognize, bind and eliminate the invading antigens.
A discussion of the molecular biology of the immune system is beyond our scope here. Suffice it to say that the recombinational pathway of immunoglobulin gene rearrangements includes enzymatic activities very similar to those of transposition. In fact, the so-called RAG1 enzyme active in immunoglobulin gene rearrangement is closely related to genes in a family of transposons (transib) found in invertebrates and fungi. Thus, it looks like genes of the immune system might have their origins in a transposon!