
2.4.3.3: Red and Green Algae
- Page ID
- 31929

Learning Objectives
- Distinguish between different groups of algae using life cycle, morphological features, and cellular composition.
- Connect adaptations in the red and green algae to habitat characteristics and ecology.
- Identify structures and phases in the Polysiphonia and Spirogyra life cycles; know the ploidy of these structures.
Red algae and green algae are included in the supergroup Archaeplastida. It was from a common ancestor of these protists that the land plants evolved, since their closest relatives are found in this group. Molecular evidence supports that all Archaeplastida are descendants of an endosymbiotic relationship between a heterotrophic protist and a cyanobacterium. This primary endosymbiosis resulted in the chloroplasts of all photosynthetic eukaryotes discussed in this book! The red and green algae include unicellular, multicellular, and colonial forms.
Glaucophyta
Glaucophytes are unicellular, phototrophic eukaryotics found in freshwater ecosystems. Glaucophytes are likely an ancestral branch in the Archaeplastida and are often used as evidence of the cyanobacterial origin of chloroplasts. They have chloroplast-like organelles, called cyanelles or muroplasts, that have peptidoglycan between the two membranes. They have the same pigments as cyanobacteria and red algae: chlorophyll a and phycobilins. Much like the cyanobacteria, they appear blue-green due to the interaction of chlorophyll and phycocyanins (glauco- comes from the Greek glaukos meaning blue-green). Non-motile cells have a rigid cell wall composed of cellulose. Motile cells have two whiplash flagella. Sexual reproduction has yet to be documented in this group.
Rhodophyta
Red algae descended from the same endosymbiotic event as the Glaucophyta. The red algae are almost exclusively marine. Some are unicellular but most are multicellular. Approximately 6,000 species have been identified. They have true chloroplasts with two membranes (no remnant peptidoglycan) containing chlorophyll a. Like the cyanobacteria, they use phycobilins as antenna pigments - phycoerythrin (which makes them red) and phycocyanin. Red pigment allows the red algae to photosynthesize at deeper depths than the green or brown algae, harnessing more of the blue light waves that penetrate deeper into the water column. Unlike green algae and plants, red algae store carbohydrates as Floridean starch in the cytosol. Some are used as food in coastal regions of Asia. Agar, the base for culturing bacteria and other microorganisms, is extracted from a red alga.
Selection Pressures and Drivers
An important aspect of understanding the life history traits of the Rhodophyta is understanding the challenges of living in a marine environment.
- Access to sunlight: Most colors of light cannot penetrate into deeper water, as they are scattered by water molecules. The wavelengths of light that reach deepest into the ocean are blue and green. Many fish that live in the deep ocean are red. Because red light does not penetrate to the depths where they live, this makes them virtually undetectable by sight. Remember, we see things because of the light that bounces off of them. Red pigments reflect red light, so no red light, no reflected light. Red algae are using a similar strategy--absorb the wavelengths of light that are not red--with a different goal: to use that absorbed light to make food. The phycoerythrin in their chloroplasts reflects red light, giving them a red appearance, and absorbs the blue light that is able to penetrate to deeper areas in the water column.
- Fertilization: The ocean is an expansive environment, often with large areas of open space between populations of organisms. In this environment, successful fertilization of an egg by a nonmotile sperm--red algae lack flagella--presents a challenge. Having multicellular haploid and diploid phases provides red algae more opportunities to produce gametes and spores. A diploid stage that clones the zygote, the carposporophyte, provides more opportunities to do meiosis from each fertilization event.
- Salinity: Marine environments are relatively high in salinity. A possible adaptation for this is to have sulfated polysaccharides in the cell wall, such as the galactans present in Rhodophyta. This is a strategy present in (potentially all) marine algae and is inferred to be an adaptation for salinity-tolerance. See this open-access article for further information.

Morphology
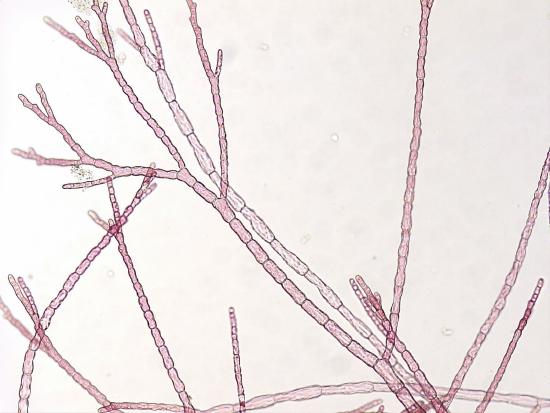
Red algae have a diverse range of morphologies. Unicellular forms may live solitarily or as colonies but, unlike other members of the Archaeplastida, lack flagella. Flagella are absent from the Rhodophyta, lost at some point in their evolutionary history. Multicellular forms can be filamentous, leafy, sheet-like, coralloid, or even crust-like (some examples in Figure \(\PageIndex{4}\) and Figure \(\PageIndex{5}\)). The strange coralline red algae have calcerous deposits in the cell walls that make the thallus hard, like a coral. These can take a variety of forms and are able to live at depths other algae cannot (over 500 feet deep for some!).



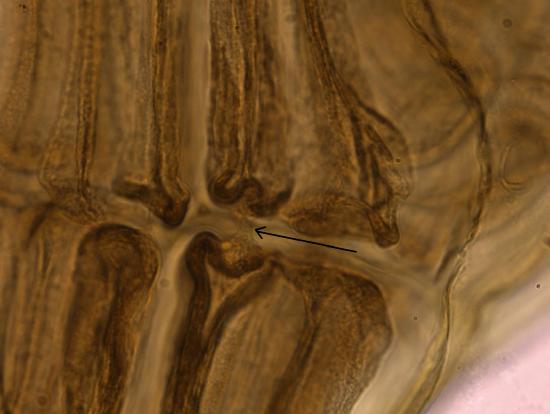
Cells of multicellular species are connected via incomplete cytokinesis, resulting in pit connections (Figure \(\PageIndex{6}\)).
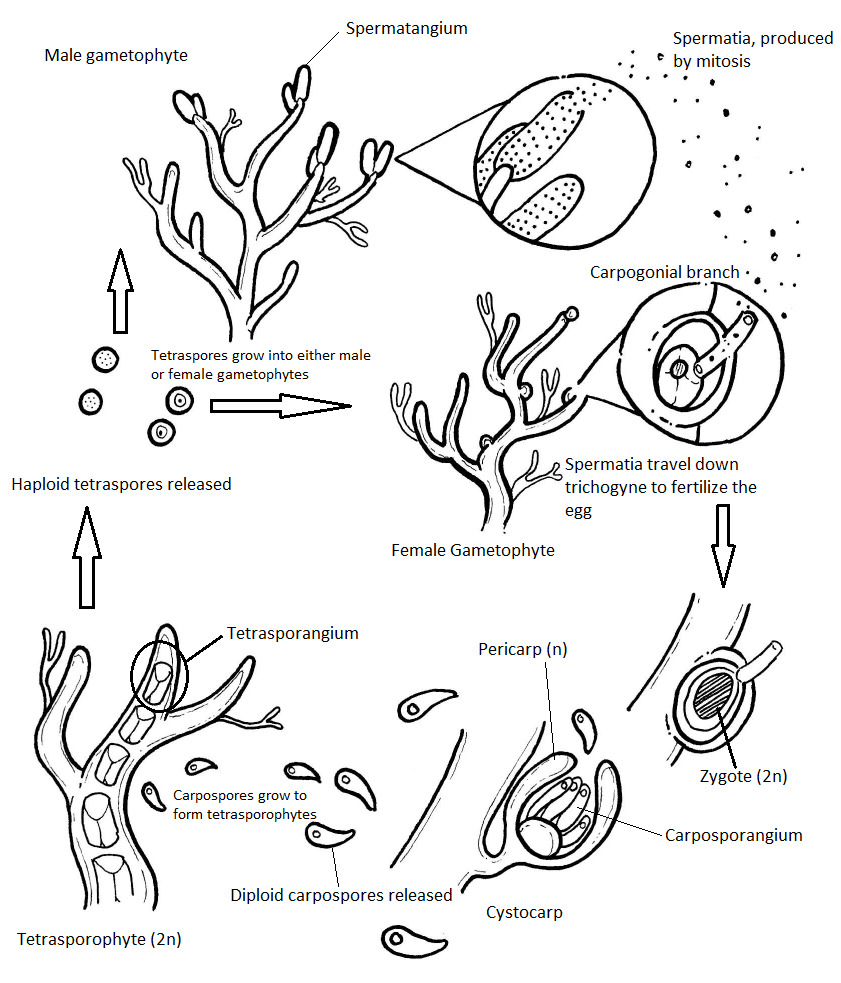
Polysiphonia Life Cycle
Red algae have a haplodiplontic (alternation of generations) life cycle that has an extra diploid stage: the carposporophyte. Polysiphonia is the model organism for the Rhodophyta life cycle. The gametophytes of Polysiphonia are isomorphic (iso- meaning same, morph- meaning form), meaning they have the same basic morphology. Any difference you see in coloration of the images in this section is due to staining. They would all appear a deep red color in an unstained slide.

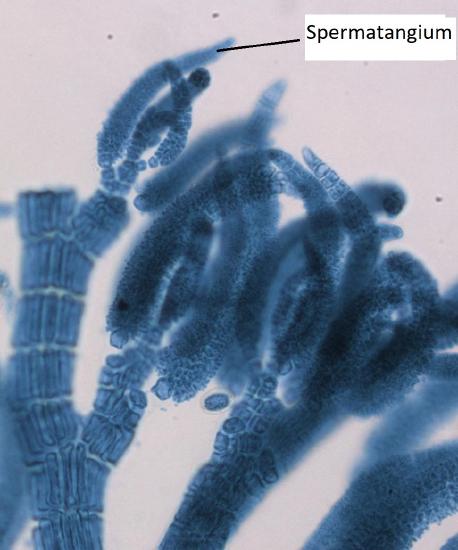
Male Gametophyte
The male gametophyte has elongated structures that emerge from the tips of the thallus branches. These are spermatangia, where spermatia are produced by mitosis.
Female Gametophyte and Carposporophyte
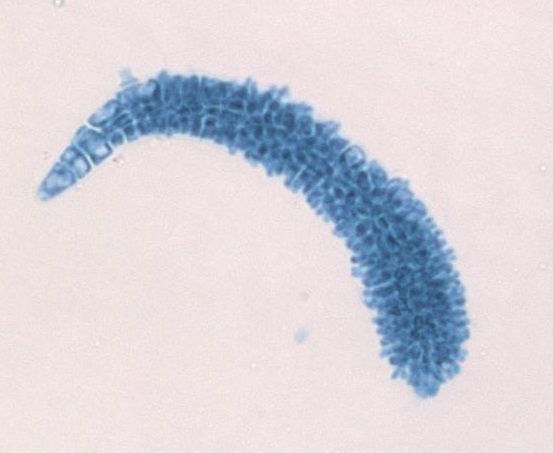
The female gametophyte produces an egg that is contained within a structure called the carpogonium. This structure has a long, thin projection called a trichogyne (trich- meaning hair, -gyne meaning female). During fertilization, a spermatium fuses with the trichogyne and the nucleus of the spermatium travels down the tube to the egg. When the nucleus of the spermatium fuses with the egg, a zygote is produced. This zygote is retained and nourished by the female gametophyte as it grows.
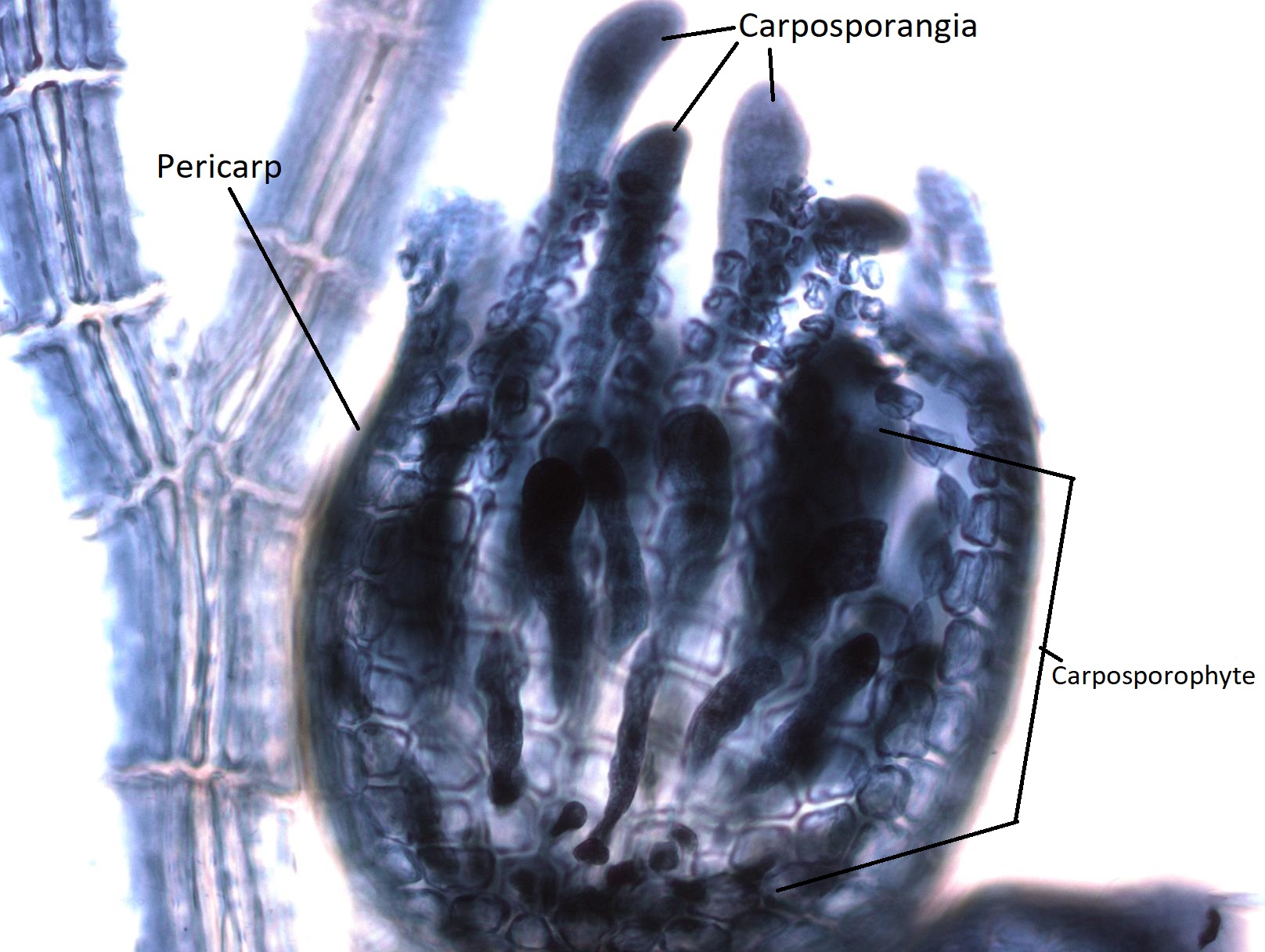
The globose structures you see growing from the female gametophyte thallus are called cystocarps. A cystocarp is composed of both female gametophyte tissue (n) and carposporophyte tissue (2n). The outer layer of the cystocarp, the pericarp (peri- meaning around) is derived from the female gametophyte and is haploid. The interior of the cystocarp consists of the carposporophyte, which is diploid, and produces structures called carposporangia, inside of which it produces carpospores by mitosis. All of these--carposporophyte, carposporangia, and carpospores--are diploid.
Tetrasporophyte
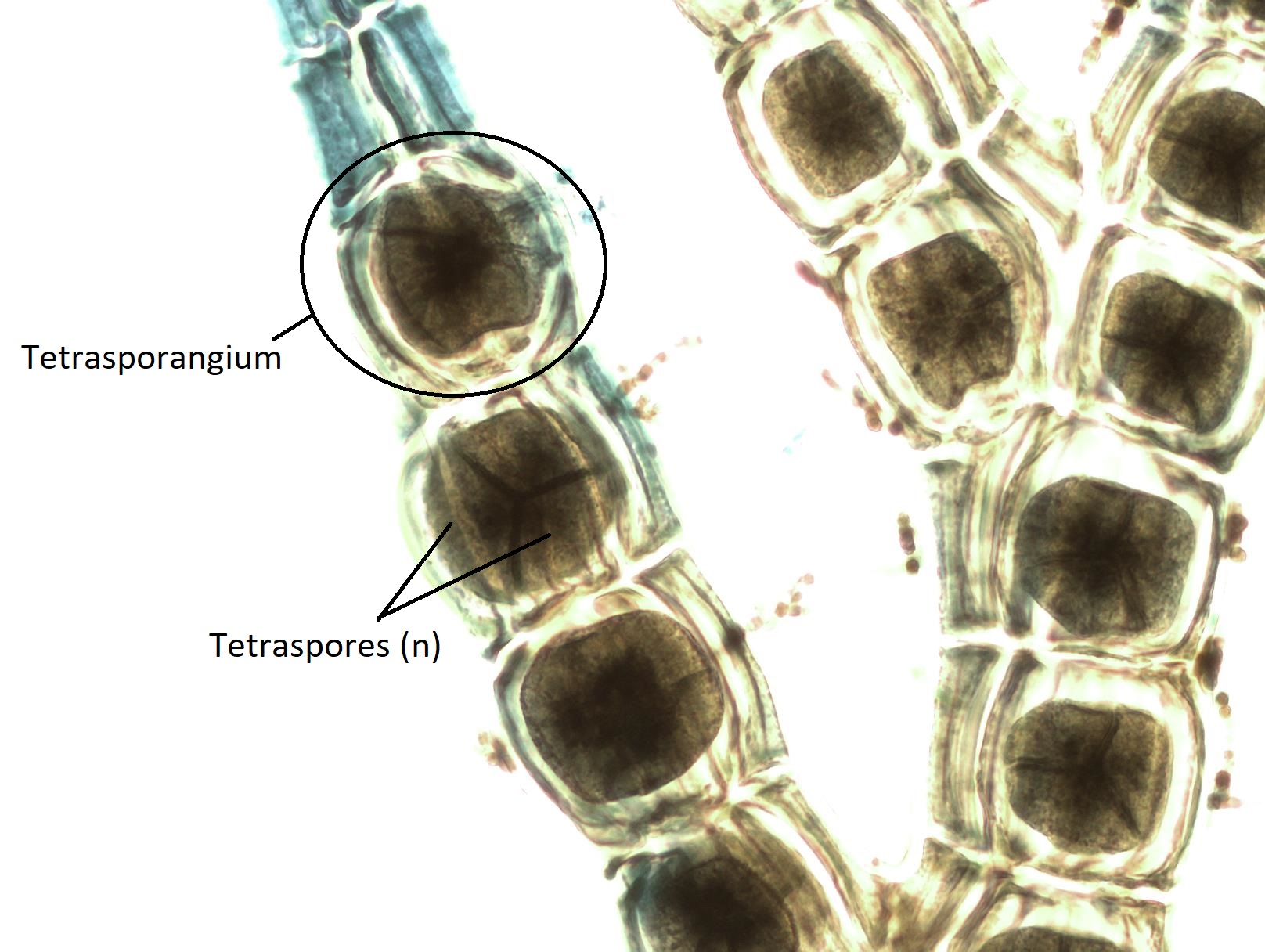
The diploid carpospores are released into the ocean waters, where they will be carried on currents to another location. If a carpospore lands in an appropriate environment, it will grow by mitosis into a tetrasporophyte (2n). The tetrasporophyte produces tetrasporangia (2n) within the branches of the thallus. Each tetrasporangium produces four unique, haploid tetraspores by meiosis. Tetraspores (n) are released and will grow by mitosis into either male or female gametophytes, completing the life cycle.
Full Life Cycle Diagram
Summary of Characteristics for Red Algae
- Morphology: Unicellular to multicellular, no flagellated stages. Cells of multicellular species are connected via incomplete cytokinesis, resulting in pit connections.
- Cell wall composition: Cellulose and galactans
- Chloroplasts: 2 membranes, pigments are chlorophyll a and phycobilins (primarily phycoerythrin, providing their red color)
- Storage carbohydrate: Floridean starch
- Life cycle: Alternation of generations with an extra diploid stage, the carposporophyte
- Ecology: Primarily marine (97% of species)
Green Algae
The most abundant group of algae is the green algae. The nature of the evolutionary relationships between the green algae are still up for debate. As of 2019, genetic data supports splitting the green algae into two major lineages: chlorophytes and streptophytes. The streptophytes include several lineages of green algae (such as the charophytes) and all land plants. Streptophytes and chlorophytes represent a monophyletic group called Viridiplantae (literally “green plants”). The green algae exhibit similar features to the land plants, particularly in terms of chloroplast structure. They have chlorophyll a and b, have lost phycobilins but gained carotenoids, and store carbohydrates as starch inside plastids. Although some of the multicellular forms are large, they never develop more than a few types of differentiated cells and their fertilized eggs do not develop into an embryo.
Green algae are an important source of food for many aquatic animals. When lakes and ponds are "fertilized" with phosphates and nitrates (e.g., from sewage and the runoff from fertilized fields and lawns), green algae often form extensive algal "blooms". Members of this group can be found in freshwater and marine habitats, and many have adapted to life on land, either inside of lichens or free-living (see Figure \(\PageIndex{12}\)).

Selection Pressures and Drivers
- Sun Damage. Green algae represent a diverse group of organisms with diverse life history traits, many of which are shared with land plants. The development of carotenoids-- yellow, orange, and red pigments that act in both light harvesting and sun protection--offers this group increased access to sunlight while simultaneously protecting against UV damage. UV rays do not penetrate very far into the water column, so organisms moving into shallower waters or terrestrial environments would need to deal with this new challenge. Many terrestrial species of green algae appear orange, rather than green, due to the production of large amounts of carotenoids.
Morphology
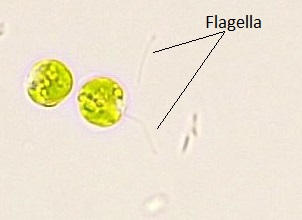
These algae exhibit great diversity of form and function. Similar to red algae, green algae can be unicellular or multicellular. Many unicellular species form colonies and some green algae exist as large, multinucleate, single cells. Green algae primarily inhabit freshwater and damp soil, and are a common component of plankton. Chlamydomonas is a simple, unicellular chlorophyte with a pear-shaped morphology and two opposing, anterior flagella that guide it toward light sensed by its eyespot (Figure \(\PageIndex{13}\)). More complex species exhibit haploid gametes and spores that resemble Chlamydomonas.
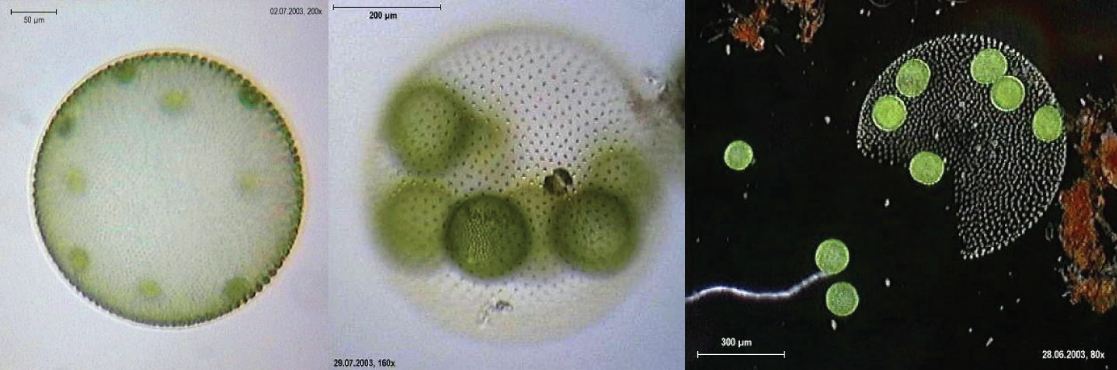
The alga Volvox is one of a colonial organism, which behaves in some ways like a collection of individual cells, but in other ways like the specialized cells of a multicellular organism (Figure \(\PageIndex{14}\)). Volvox colonies contain 500 to 60,000 cells, each with two flagella, contained within a hollow, spherical matrix composed of a gelatinous glycoprotein secretion. Individual Volvox cells move in a coordinated fashion and are interconnected by cytoplasmic bridges. Only a few of the cells reproduce to create daughter colonies, an example of basic cell specialization in this organism.
Volvox can reproduce both asexually and sexually. In asexual reproduction, the gonidia develop into new organisms that break out of the parent (which then dies). In sexual reproduction, the presence of an inducing chemical causes the following:
- The gonidia of the males to develop into clusters of sperm.
- The gonidia of the females to develop into new spheres each of whose own gonidia develops into a pair of eggs.
- The sperm break out of the male parent and swim to the female where they fertilize her eggs.
- The zygotes form a resting stage that enables Volvox to survive harsh conditions (Figure \(\PageIndex{15}\)).
Video \(\PageIndex{1}\): This video shows how sexual reproduction occurs in the colonial green alga Volvox. Sourced from YouTube.
The genome of Volvox carteri consists of 14,560 protein-encoding genes - only 4 more genes than in the single-celled Chlamydomonas reinhardtii! Most of its genes are also found in Chlamydomonas. The few that are not encode the proteins needed to form the massive extracellular matrix of Volvox.
Species in the genus Caulerpa exhibit flattened fern-like foliage and can reach lengths of 3 meters (Figure \(\PageIndex{16}\)). Caulerpa species undergo nuclear division, but their cells do not complete cytokinesis, remaining instead as massive and elaborate single cells.

True multicellular organisms, such as the sea lettuce, Ulva, are also represented among the green algae (Figure \(\PageIndex{17}\) and Figure \(\PageIndex{18}\)).


Spirogyra Life Cycle
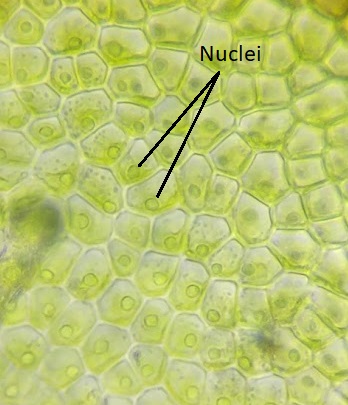
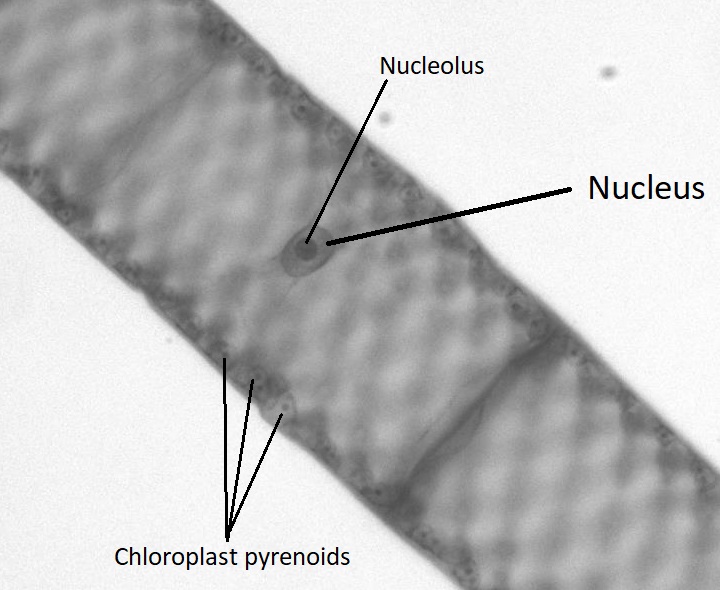
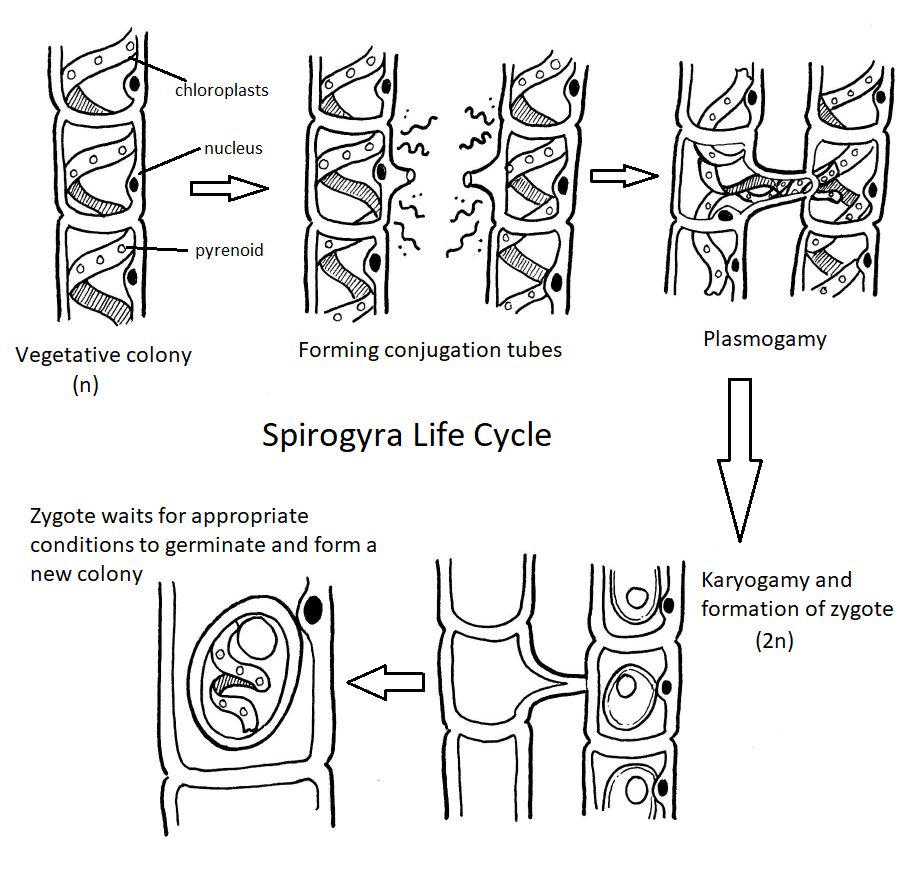
Though green algae display a diversity of life cycles, many have a haplontic life cycle. A model organism for the green algae is Spirogyra (Figure \(\PageIndex{19}\)). Spirogyra is a unicellular green algae that grows in long, filamentous colonies, making it appear to be a multicellular organism. Even though it is technically unicellular, its colonial nature allows us to classify its life cycle as haplontic. In the haploid vegetative cells of the colony, the chloroplasts are arranged in spirals, containing darkened regions called pyrenoids where carbon fixation happens. Each haploid cell in the filament is an individual, which makes sexual reproduction between colonies an interesting process.
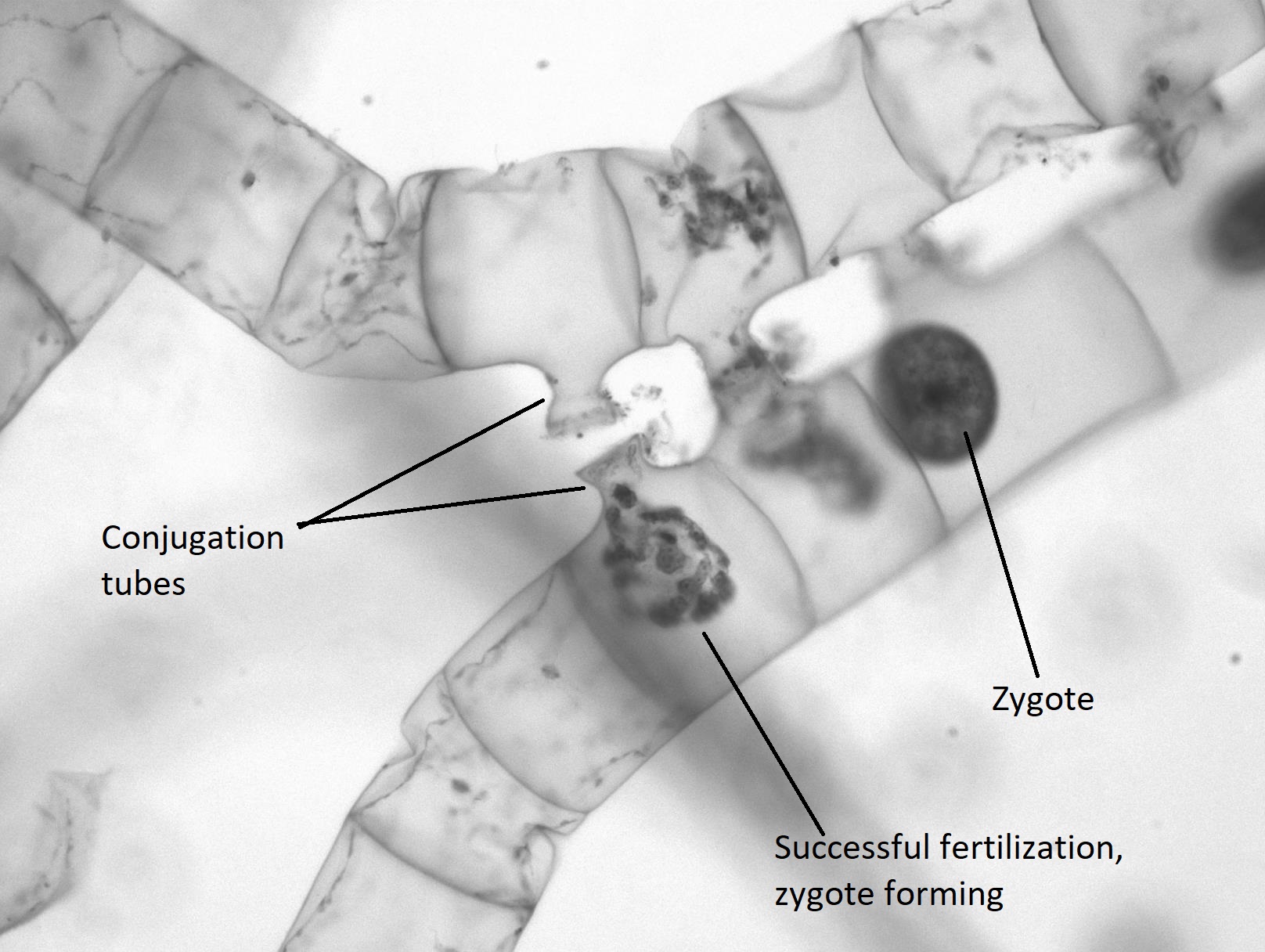
When two colonies of Spirogyra meet that are of a complementary mating type (+/-), sexual reproduction occurs. The two colonies align, each cell across from a complementary cell on the other filament. A conjugation tube extends from each cell in one colony (Figure \(\PageIndex{20}\)), inducing formation of a tube on the cells in the other colony. The conjugation tubes from each colony fuse together.

The contents of one cell will move through the conjugation tube and fuse with the contents of the complementary cell, resulting in a diploid zygote (Figure \(\PageIndex{21}\)). The zygote appears as a large, egg-like structure contained within the complementary cell. It has a thick wall that provides resistance to desiccation and cold, allowing colonies of Spirogyra to overwinter, when needed. The other colony is now a filament of empty cells that will be broken down by some decomposer. When conditions are right, the zygote undergoes meiosis to produce another vegetative colony of haploid cells.
Full Life Cycle Diagram
Summary of Characteristics for Green Algae
- Morphology: Unicellular to multicellular; two whiplash flagella on motile cells
- Cell wall composition: Cellulose
- Chloroplasts: 2 membranes, pigments are chlorophyll a, chlorophyll b, and carotenoids
- Storage carbohydrate: Starch
- Life cycle: Varies, but primarily haplontic. Some marine species have alternation of generations.
- Ecology: Freshwater, marine, and terrestrial species
Summary
Glaucophytes, red algae, and green algae are part of the Archaeplastida. These organisms are descended from the same primary endosymbiosis event. Glaucophytes are thought to be one of the earliest lineages to diverge due to the presence of remnant peptidoglycan between the membranes of its chloroplast-like cyanelles. Unsurprisingly, glaucophytes and red algae share the same pigments as Cyanobacteria.
Red algae (phylum Rhodophyta) are united by several synapomorphies (shared derived characteristics). They lack flagella, have pit connections between cells, and store carbohydrates as Floridean starch. The sulfated galactans in their cell walls allows them increased fitness in marine environments, while the pigment phycoerythrin allows them to photosynthesize deeper in the water column. They have an alternation of generations life cycle with an extra diploid phase, the carposporophyte, that clones the zygote. These characteristics can be connected to the environmental stressors presented by the marine habitats most red algae are found in.
Green algae represent several distinct lineages. Like plants, they store carbohydrates as starch within their plastids and have the pigments chlorophyll a and b, as well as carotenoids. Organisms in this group have haplontic (e.g. Spirogyra) or haplodiplontic (e.g. Ulva) life cycles. Many green algae are unicellular, forming complex colonies. Green algae can be found in marine, freshwater, and terrestrial environments (including within lichens!).
Attributions
Curated and authored by Maria Morrow, CC BY-NC, using the following sources:
- 19.1.2 Protists, 19.1.4 Volvox, and 19.1.5 Diversity and Evolutionary Relationships of the Plants from Biology by John. W. Kimball (licensed CC-BY)
- 23.3 Groups of Protists from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.