
21.2: Biosynthesis of Triacylglycerols
- Page ID
- 15174


by William (Bill) W. Christie and Henry Jakubowski.
This section is an abbreviated and modified version of material from the Lipid Web, an introduction to the chemistry and biochemistry of individual lipid classes, written by William Christie.
Introduction
All eukaryotic organisms and even a few prokaryotes can synthesize triacylglycerols, and in animals, many cell types and organs have this ability, but the liver, intestines, and adipose tissue are most active with most of the body stores in the last of these. Within all cell types, even those of the brain, triacylglycerols are stored as cytoplasmic 'lipid droplets' enclosed by a monolayer of phospholipids and hydrophobic proteins such as the perilipins in adipose tissue or oleosins in seeds. These lipid droplets are now treated as distinctive organelles, with their own characteristic metabolic pathways and associated enzymes - no longer boring blobs of fat. However, they are not unique to animals and plants, as Mycobacteria and yeasts have similar lipid inclusions.
The lipid serves as a store of fatty acids for energy, which can be released rapidly on demand, and as a reserve of fatty acids for structural purposes or as precursors for eicosanoids. In addition, lipid droplets serve as a protective agency in cells to sequester any excess of biologically active and potentially harmful lipids such as free fatty acids, oxylipins, diacylglycerols, cholesterol (as cholesterol esters), retinol esters, and coenzyme A esters.
While triacylglycerols are essential for normal physiology, an excessive accumulation in human adipose tissue and other organs results in obesity and other health problems, including insulin resistance, steatohepatitis, and cardiomyopathy. Accordingly, there is considerable pharmaceutical interest in drugs that affect triacylglycerol biosynthesis and metabolism.
Biosynthesis of Triacylglycerols
Three main pathways for triacylglycerol biosynthesis include the sn-glycerol-3-phosphate and dihydroxyacetone phosphate pathways, which predominate in liver and adipose tissue, and a monoacylglycerol pathway in the intestines. In maturing plant seeds and some animal tissues, a fourth pathway has been recognized in which a diacylglycerol transferase is involved.
The most important route to triacylglycerols is the sn-glycerol-3-phosphate or Kennedy pathway, first described by Eugene Kennedy and colleagues in the 1950s, from which more than 90% of liver triacylglycerols are produced. Figure \(\PageIndex{1}\) shows the pathway from glycerol-3-phosphate to triacylglycerols (the Kennedy pathway) and some preceding reactions that generate glycerol-3-phosphate.

In this pathway, the main source of the glycerol backbone has long been believed to be sn-glycerol-3-phosphate produced by the catabolism of glucose (glycolysis) or to a lesser extent by the action of the enzyme glycerol kinase on free glycerol. However, there is increasing evidence that a significant proportion of glycerol is produced de novo by a process known as glyceroneogenesis via pyruvate. Indeed, this may be the main source in adipose tissue.
Subsequent reactions occur primarily in or at the endoplasmic reticulum. First, the precursor sn-glycerol-3-phosphate is esterified by a fatty acid coenzyme A ester in a reaction catalyzed by a glycerol-3-phosphate acyltransferase (GPAT) at position sn-1 to form lysophosphatidic acid, and this is in turn acylated by an acylglycerophosphate acyltransferase (AGPAT) in position sn-2 to form a key intermediate in the biosynthesis of all glycerolipids - phosphatidic acid. Numerous isoforms of these enzymes are known; they are expressed with specific tissue and membrane distributions, and they are regulated in different ways.
Let's look at some of the enzymes involved in some key reactions in the synthesis of triacylglycerols.
GPAT Enzymes
Glycerol-3-phosphate acyltransferase (GPAT) catalyzes the first step in the pathway in most tissue. From an enzymatic perspective, it can be considered rate-limiting for the pathways since its specific activity is slow. In mammals, there are four isoforms (GPAT1 in the outer membrane of the mitochondria, GPAT2 also in the mitochondria, and GPAT3 and 4, both in the ER. The activity of GPAT1 is activated by insulin, presumably through phosphorylation) and inhibited by AMP-activated protein kinase (AMPK).
Fatty acyl-CoA in the outer membrane could be used for TAG synthesis through the Kennedy pathway in times of energy abundance or imported into the matrix through the carnitine-acyl-CoA cycle in times of energy need. Overexpression of GPAT1 in liver cells increases TAG synthesis and decreases β-oxidation. GPAT1 deficient mice in contrast have higher levels of β-oxidation.
Figure \(\PageIndex{2}\) shows an interactive iCn3D model of the AlphaFold predicted structure of human mitochondrial glycerol-3-phosphate acyltransferase 1 - GPAT1 (Q9HCL2).
.png?revision=1&size=bestfit&width=376&height=359)
 Figure \(\PageIndex{2}\): AlphaFold predicted structure ofhuman mitochondrial glycerol-3-phosphate acyltransferase 1 - GPAT1 (Q9HCL2). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...kuvBTPo5mNK8p9
Figure \(\PageIndex{2}\): AlphaFold predicted structure ofhuman mitochondrial glycerol-3-phosphate acyltransferase 1 - GPAT1 (Q9HCL2). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...kuvBTPo5mNK8p9
Four conserved motifs involved in catalysis (red and cyan) and binding (orange and yellow) are shown. Significant sections are still disordered in the model. Two highly conserved amino acids, His 230 and Asp 235 in the red catalytic motif are shown in CPK-colored sticks and labeled. The black helices are predicted outer mitochondrial membrane transmembrane helices. Both the N- and C-terminal amino acids project into the cytoplasm.
Phosphatidic acid phosphohydrolases (PAPs or ‘phosphatidate phosphatases’ or ‘lipid phosphate phosphatases’).
PAPs are also important as they produce sn-1,2-diacylglycerols as essential intermediates in the biosynthesis not only of triacylglycerols but also of phosphatidylcholine and phosphatidylethanolamine (and of monogalactosyldiacylglycerols in plants). This is a key branch-point in lipid biosynthesis as it may dictate the flow of lipids for storage or membrane biogenesis.
Much of this phosphatase activity leading to triacylglycerol biosynthesis in animals reside in three related cytoplasmic proteins, termed lipins, i.e., lipin-1, lipin-2 and lipin-3, which have tissue-specific roles in glycerolipid synthesis. Unusually, these were characterized and named before the nature of their enzymatic activities was determined. Each of the lipins appears to have distinctive expression and functions, but lipin-1 (PAP1) in three isoforms (designated 1α, 1β, and 1γ) accounts for most of the PAP activity in adipose tissue and skeletal muscle in humans. Lipin 2 is the most abundant lipin in the liver but is also expressed substantially in the small intestine, macrophages, and some regions of the brain, while lipin 3 activity overlaps with that of lipin 1 and lipin 2 and is found in the gastrointestinal tract and liver. Lipins are cytosolic enzymes but associate transiently with membranes to access their substrate, i.e., they are translocated to the endoplasmic reticulum in response to elevated levels of fatty acids within cells, although they do not have trans-membrane domains. Lipin-1 activity requires Mg2+ ions and is inhibited by N-ethylmaleimide, whereas the membrane-bound activity responsible for synthesizing diacylglycerols as a phospholipid intermediate is independent of Mg2+ concentration and is not sensitive to the inhibitor.
Perhaps surprisingly, lipin-1 has a dual role in that it operates in collaboration with known nuclear receptors as a transcriptional coactivator to modulate lipid metabolism (lipin 1α) while lipin 1β is associated with the induction of lipogenic genes such as fatty acid synthase, stearoyl-CoA desaturase, and DGAT. They can have profound effects on signaling in a variety of cell types. Abnormalities in lipin-1 expression are known to be involved in some human disease states that may lead to metabolic syndrome and inflammatory disorders. Lipin 2 is a similar phosphatidate phosphohydrolase, which is regulated dynamically by fasting and obesity (in mice).
PAP depend on divalent Mg2+ ions and are sensitive to N-ethylmaleimide. Lipin has a N-Lip (N-terminal) and C-Lip (C-terminal) domain. The crystal structure of lipin with a 250 amino acid regions between the two domains removed has been solved. The domain structures of humans, the yeast Saccharomyces cerevisiae (SC) and its truncated form (Tt-Pah2), and the structure of the Tt-Pah2 truncated fragment of the enzyme from Tetrahymena thermophila, are shown in Figure \(\PageIndex{3}\).

Panel (a) shows the PAP-catalyzed dephosphorylation reaction.
Panel (b) shows the domain architecture of PAPs drawn to scale. The positions of the nuclear localization signal (NLS), conserved Trp-motif (purple W), catalytic DxDxT motif, and fatty liver dystrophy (fld2J) mutation are indicated.
Panel (c) shows the the wild-type (WT) Tt Pah2 is catalytically active and the D146A mutant of the DxDxT motif eliminates activity.
Panel (d) shows the overall structure of Tt Pah2, which contains an immunoglobulin-like (Ig-like) domain and a HAD-like catalytic domain. The Ig-like domain is formed by the N-Lip (cyan) and C-Lip (pink) regions connected by a short linker (gray loop) that would be replaced by the extended 500-residue linker in human lipins and a 250-residue linker in Sc Pah1. A calcium ion (Ca2+, yellow sphere) is bound in the active site of the HAD-like catalytic domain. The top view (right) of the enzyme shows the N-Lip co-folding with the C-Lip to form the Ig-like domain.
Figure \(\PageIndex{4}\)s shows a closeup of the active site of Tt Pah2 and a mechanism for the hydrolytic removal of the phosphate on DAG.

Figure \(\PageIndex{5}\) shows an interactive iCn3D model of the Tetrahymena Thermophila lipin phosphatidic acid phosphatase with magnesium (6TZZ).
.png?revision=1&size=bestfit&width=397&height=278)
Figure \(\PageIndex{5}\): Tetrahymena Thermophila lipin phosphatidic acid phosphatase with magnesium (6TZZ). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...GTusCCDP5eRrt7
The N-terminal domain is shown in lavender and the C-terminal domain is in gray. Key amino acids in the active site are shown in CPK-colored sticks and labeled.
Diacylglycerol acyltransferases (DGAT)
In the final step in this pathway, the 1,2-diacyl-sn-glycerol intermediate is acylated by diacylglycerol acyltransferases (DGAT), which can utilize a wide range of fatty acyl-CoA esters to form the triacyl-sn-glycerol. There are two DGAT enzymes, which are structurally and functionally distinct. In animals, DGAT1 is located mainly in the endoplasmic reticulum and is expressed in skeletal muscle, skin, and intestine, with lower levels of expression in the liver and adipose tissue. It is believed to have dual topology contributing to triacylglycerol synthesis on both sides of the membrane of the endoplasmic reticulum but esterifying only pre-formed fatty acids of exogenous origin. Perhaps surprisingly, DGAT1 is the only one present in the epithelial cells that synthesize milk fat in the mammary gland. Also, DGAT1 can utilize a wider range of substrates, including monoacylglycerols, long-chain alcohols (for wax synthesis), and retinol, and it is reported to have an important role in protecting the endoplasmic reticulum from the lipotoxic effects of high-fat diets. Orthologs of this enzyme are present in most eukaryotes, other than yeasts, and they are especially important in plants.
Figure \(\PageIndex{6}\) shows an interactive iCn3D model of Human Diacylglycerol Acyltransferase 1 in complex with oleoyl-CoA (6VP0)
.png?revision=1&size=bestfit&width=469&height=260)
Figure \(\PageIndex{6}\): Human Diacylglycerol Acyltransferase 1 in complex with oleoyl-CoA (6VP0) (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...adyWg9fy2hh8r8
The red layer is the extracellular leaflet and the blue is the intracellular leaflet. The protein is a dimer with one subunit shown in gray and one in light. A substrate, oleoyl-CoA, is shown in spacefill with CPK colors in both monomers. The acyl chain with a clear kink induced by the double bond in the oleoyl acyl group is clearly in the hydrophobic interior of the bilayer. The gray monomer shows the atoms within 4 Å from the oleoly-CoA is shown as surface color-coded by hydrophobicity with the green most hydrophobic. The active site His 415 and Met 434 which hydrogen bonds to it are shown in the light cyan subunit in sticks, CPK colors, and labeled. Three other conserved polar side chains, Asn378, Gln437, and Gln465, which are in the active site region, are also shown.
Each monomer in the homodimer has 9 transmembrane helices with 8 forming a unique fold called the MBOAT which encloses a chamber with catalytic side chains. The substrates, fatty acyl-CoA, and DAG have different entrances to the chamber with the acyl-CoA occupying a cytosolic tunnel. The cytosolic face interacts with the CoA and the acyl chain extends into the ER membrane. The channel appears bent which probably allows the acylation of DAGs and long-chain alcohols, but not cholesterol, a rigid, planar molecule that is acylated to form an ester by a different enzyme, acyl CoA:cholesterol acyltransferases. Hence it appears that DAG, the acyl-acceptor, binds in a hydrophobic tunnel from within the bilayer and the acyl-CoA binds in another tunnel, allowing the transfer of the acyl group.
DGAT2 is the main form of the enzyme in hepatocytes and adipocytes (lipid droplets), although it is expressed much more widely in tissues. It is associated with distinct regions of the endoplasmic reticulum, at the surface of lipid droplets, and in mitochondria, and it esterifies fatty acids of both endogenous and exogenous origin. DGAT2 is believed to have a targeting domain that enables it to tether between the endoplasmic reticulum and lipid droplet thereby channeling triacylglycerols from the synthesis site in the endoplasmic reticulum to the nascent lipid droplet, where they accumulate and lead to the expansion of the latter (see below). Both enzymes are important modulators of energy metabolism, although DGAT2 appears to be especially important in controlling the homeostasis of triacylglycerols in vivo. As the glycerol-3-phosphate acyltransferase (GPAT) has the lowest specific activity of these enzymes, this step may be the rate-limiting one. However, DGATs are the dedicated triacylglycerol-forming enzymes, and they are seen as the best target for pharmaceutical intervention in obesity and attendant ailments; clinical studies of DGAT1 inhibitors are at an early stage.
In a second pathway for triacylglycerol biosynthesis, dihydroxyacetone-phosphate in peroxisomes or endoplasmic reticulum can be acylated with fatty acid CoA esters by a specific acyltransferase to form 1-acyl dihydroxyacetone-phosphate, which is reduced by dihydroxyacetone-phosphate oxidoreductase to lysophosphatidic acid; this can then enter the pathway above to triacylglycerols. The precursor dihydroxyacetone-phosphate is important also as part of the biosynthetic route to plasmalogens, and neutral plasmalogens can be significant components of cytoplasmic droplets in many mammalian cells types but not in adipose tissue.
Figure \(\PageIndex{7}\): Biosynthesis of triacylglycerols from dihydroxyacetone-phosphate

In prokaryotes, the glycerol-3-phosphate pathway of triacylglycerol biosynthesis only occurs, but in yeast, both glycerol-3-phosphate and dihydroxyacetone-phosphate can be the primary precursors and synthesis takes place in cytoplasmic lipid droplets and the endoplasmic reticulum. In plants, the glycerol-3-phosphate pathway is most important.
In the enterocytes of the intestines after a meal, up to 75% of the triacylglycerols are formed via a monoacylglycerol pathway. In this, 2-monoacyl-sn-glycerols and free fatty acids released from dietary triacylglycerols by the action of pancreatic lipase within the intestines (see below) are taken up by the enterocytes. There, the monoacylglycerols are first acylated by an acyl-coenzyme A:monoacylglycerol acyltransferase with the formation of sn-1,2-diacylglycerols mainly as the first intermediate in the process, though some sn-2,3-diacylglycerols (~10%) are produced. In addition, 1-monoacylglycerols can be synthesized by acylation of glycerol for further acylation. There are three isoforms of the monoacylglycerol acyltransferase in humans of which MGAT2 is most active in the intestines, but also in the liver where an appreciable proportion of the triacylglycerols are formed by the monoacylglycerol pathway, while MGAT1 functions in adipose tissue; the role of MGAT3 is not clear. Finally, the acyl-coenzyme A:diacylglycerol acyltransferase (DGAT1) reacts with the sn-1,2-diacylglycerols (not the sn-2,3 form) to produce triacylglycerols (DGAT1 can also acylate monoacylglycerols). Figure \(\PageIndex{8}\) shows the biosynthesis of triacylglycerols from monoacylglycerols.

Other pathways: In a fourth biosynthetic pathway, which is less well known, triacylglycerols are synthesized by a transacylation reaction between two racemic diacylglycerols that are independent of acyl-CoA. The reaction was first detected in the endoplasmic reticulum of intestinal micro villus cells and is catalyzed by a diacylglycerol transacylase. Both diacylglycerol enantiomers participate in the reaction with equal facility to transfer a fatty acyl group with the formation of triacylglycerols and a 2-monoacyl-sn-glycerol. A similar reaction has been observed in seed oils. Figure \(\PageIndex{9}\): Biosynthesis of triacylglycerols from diacylglycerol transacylases

It has been suggested that this enzyme may function in remodeling triacylglycerols post-synthesis, especially in oil seeds, and it may be involved in similar processes in the liver and adipose tissue, where extensive hydrolysis/re-esterification is known to occur. There is evidence for selectivity in the biosynthesis of different molecular species in a variety of tissues and organisms, which may be a consequence of the varying biosynthetic pathways. Also in adipose tissue, fatty acids synthesized de novo are utilized in different ways from those from external sources in that they enter positions sn-1 and 2 predominantly, while a high proportion of the oleic acid synthesized in the tissue by desaturation of exogenous stearic acid is esterified to position sn-3.
Among other potential routes to the various intermediates, lysophosphatidic acid and phosphatidic acid can be synthesized in mitochondria, but they must then be transported to the endoplasmic reticulum before they enter the pathway for triacylglycerol production. 1,2‑Diacyl-sn-glycerols are also produced by the action of phospholipase C on phospholipids and can be utilized for triacylglycerol biosynthesis.
In the glycerol-3-phosphate and other pathways, the starting material is of defined stereochemistry and each of the enzymes catalyzing the various steps in the process is distinctive and can have preferences for particular fatty acids (as their coenzyme A esters) and for particular fatty acid combinations in the partially acylated intermediates. It should not be surprising, therefore, that natural triacylglycerols exist in enantiomeric forms with each position of the sn-glycerol moiety esterified by different fatty acids.
Triacylglycerol Metabolism in the Intestines, Liver, and Mammary Gland
Fat comprises up to 40% of the energy intake in the human diet in Western countries, and a high proportion of this is in the form of triacylglycerols. The process of fat digestion is begun in the stomach with acid-stable gastric or lingual lipases, the extent of which depends on species but may be important for efficient emulsification. However, this is insignificant in quantitative terms in comparison to the reaction with pancreatic lipase, which occurs in the duodenum. Entry of triacylglycerol degradation products into the duodenum stimulates the synthesis of the hormone cholecystokinin and causes the gall bladder to release bile acids, which are strong detergents and act to emulsify the hydrophobic triacylglycerols so increasing the available surface area. In turn, cholecystokinin stimulates the release of the hydrolytic enzyme pancreatic lipase together with a co-lipase, which is essential for the activity of the enzyme. Pancreatic lipase, co-lipase, bile salts, and calcium ions act together in a complex at the surface of the emulsified fat droplets to hydrolyze the triacylglycerols. The process is regiospecific and results in the release of the fatty acids from the 1 and 3 positions and the formation of 2-monoacyl-sn-glycerols as shown in Figure \(\PageIndex{10}\).

Spontaneous isomerization of the latter to 1(3)-monoacyl-sn-glycerols occurs to some extent, and these can be degraded completely by the enzyme to glycerol and free fatty acids. Other lipases hydrolyze the phospholipids and other complex lipids in foods at the same time.
This process is somewhat different in neonates and young infants, in whom pancreatic lipase is less active but is effectively replaced by lipases in breast milk and by an acid gastric lipase (pH optimum 4-6).
There is evidence that the regiospecific structure of dietary triacylglycerols affects the uptake of particular fatty acids and may influence further lipid metabolism in humans. Incorporation of palmitic acid into the position sn-2 of milk fat may be of benefit to the human infant (as a source of energy for growth and development), although it increases the atherogenic potential for adults. In addition, 2-monoacylglycerols and 2-oleoylglycerol especially have a signaling function in the intestines by activating a specific G‑protein coupled receptor GPR119, sometimes termed the ‘fat sensor’. When stimulated, this causes a reduction in food intake and body weight gain in rats and regulates glucose-stimulated insulin secretion. The free fatty acids released have a similar effect, though by a very different mechanism, via the receptor GPR40. Overall, it has become evident that triacylglycerol metabolism in the intestine has regulatory effects on the secretion of gut hormones and subsequently on systemic lipid metabolism and energy balance.
The free fatty acids and 2-monoacyl-sn-glycerols are rapidly taken up by the intestinal cells, from the distal duodenum to the jejunum, via specific carrier molecules but also by passive diffusion. A specific fatty acid binding protein prevents a potentially toxic build-up of unesterified fatty acids and targets them for triacylglycerol biosynthesis. The long-chain fatty acids are converted to the CoA esters and esterified into triacylglycerols by the monoacylglycerol pathway as described above. In contrast, short and medium-chain fatty acids (C12 and below) are absorbed in unesterified form and pass directly into the portal blood stream, where they are transported to the liver to be oxidized.
Subsequently, the triacylglycerols are incorporated into lipoprotein complexes termed chylomicrons in the enterocytes in the small intestines. In brief, these consist of a core of triacylglycerols together with some cholesterol esters that is stabilized and rendered compatible with an aqueous environment by a surface film consisting of phospholipids, free cholesterol, and one molecule of a truncated form of apoprotein B (called apo B48). A cartoon structure of a chylomicron is shown in Figure \(\PageIndex{11}\).

These particles are secreted into the lymph and thence into the plasma for transport to the peripheral tissues for storage or structural purposes. Adipose tissue in particular exports appreciable amounts of the enzyme lipoprotein lipase, which binds to the luminal membrane of endothelial cells facing into the blood, where it rapidly hydrolyses the passing triacylglycerols at the cell surface releasing free fatty acids, most of which are absorbed into the adjacent adipocytes and re-utilized for triacylglycerol synthesis within the cell.
The chylomicrons remnants eventually reach the liver, where the remaining lipids are hydrolyzed at the external membranes by a hepatic lipase and absorbed. The fatty acids within the liver can be utilized for a variety of purposes, from oxidation to the synthesis of structural lipids, but a proportion is re-converted into triacylglycerols, and some of this is stored as lipid droplets within the cytoplasm of the cells (see next section). In addition, phosphatidylcholine from the high-density lipoproteins is taken up by the liver, and a high proportion of this is eventually converted to triacylglycerols. In a healthy liver, the levels of triacylglycerols are low (<5% of the total lipids), because the rates of acquisition of fatty acid from plasma and synthesis de novo within the liver are balanced by rates of oxidation and secretion into plasma. On the other hand, excessive accumulation of storage triacylglycerols is associated with fatty liver, insulin resistance, and type 2 diabetes.
Most of the newly synthesized triacylglycerols are exported into the plasma in the form of very-low-density lipoproteins (VLDL), consisting again of a triacylglycerol and cholesterol ester core, surrounded by phospholipids and free cholesterol, together with one molecule of full-length apoprotein B (100 kDa), apoprotein C and sometimes apoprotein E. These particles in turn are transported to the peripheral tissues, where they are hydrolyzed and the free acids absorbed. Eventually, the remnants are returned to the liver.
In the mammary gland, triacylglycerols are synthesized in the endoplasmic reticulum and large lipid droplets are produced with a monolayer of phospholipids derived from this membrane. These are transported to the plasma membrane and bud off into the milk with an envelope comprised of the phospholipid membrane to form milk fat globules as food for the newborn. The process is thus very different from that involved in the secretion of triacylglycerol-rich lipoproteins from other organs.
Triacylglycerol Synthesis and Catabolism (Lipolysis) in Adipocytes and Lipid Droplets
Adipose tissue and the adipocytes are characterized by accumulations of triacylglycerols, which act as the main energy store for animals, although they also cushion and insulate the body. Large fat depots occur around internal organs such as the liver, and also subcutaneously, and each of these may react differently to metabolic constraints. Thus, triacylglycerols stored when there is a surplus of nutrients are mobilized for energy production during starvation. Adipose tissue also functions as a reserve of bioactive lipids, such as eicosanoids and lipid-soluble vitamins, and when required provides structural components, including fatty acids, cholesterol, and retinol, for membrane synthesis and repair. By buffering against fatty acid accumulation that might exceed their capacity, non-adipose cells defend themselves in this way against lipotoxicity while providing a rapid source of energy and essential metabolites by sensing and responding rapidly to changes in systemic energy balance. Brown and beige fat have special properties and are discussed below, while bone marrow adipocytes (70% of the available space) have distinctive functions also.
Similarly, within most other animal cells, including most cell types in the brain, a proportion of the fatty acids taken up from the circulation is converted to triacylglycerols as described above and incorporated into cytoplasmic lipid droplets (also termed 'fat globules', 'oil bodies', 'lipid particles' or 'adiposomes'). In adipocytes, the lipid droplets can range from up to 200 μm in diameter, while other cell types contain smaller lipid droplets of the order of 50 nm in diameter. The triacylglycerol droplets together with cholesterol esters and other neutral lipids are surrounded by a protective monolayer that includes phospholipids, cholesterol, and hydrophobic proteins. The phospholipid component of the monolayer consists mainly of phosphatidylcholine and phosphatidylethanolamine derived from cytosolic leaflets of the endoplasmic reticulum and plasma membrane. Among the proteins are many that function directly in lipid metabolism, and they include acyltransferases, lipases, perilipins, caveolins, and the Adipose Differentiation Related Protein (ADRP or adipophilin). Acting in concert with other cellular organelles, they function in many different metabolic processes facilitating coordination and communication between different organelles and acting as vital hubs of cellular metabolism.
Cytosolic lipid droplets with similar metabolic activities are found in most eukaryotic cells, including those of the fruit fly Drosophila melanogaster, and many aspects of triacylglycerol processing and regulation parallel those in humans. They are also present in the cytoplasm of some prokaryotes and in the plastids and other organelles of plants (see below).
Lipid droplet assembly
This process takes place in sub-domains of the endoplasmic reticulum, where at least one isoform of each of the enzymes of triacylglycerol biosynthesis, from acyl-CoA synthetases through to glycerol-3-phosphate acyltransferases, is located probably in a protein assembly or 'interactome'. As triacylglycerols accumulate, they reach a critical level when a spontaneous condensation or nucleation by phase separation occurs, leading to the formation of an oil blister within the hydrophobic bilayer region so attracting perilipins and other proteins that allow lipid droplets to grow further in patches of the membrane as lens-like swellings between the two membrane leaflets.
The protein seipin stabilizes the nascent droplets with minimal disruption to the membrane and enables them to mature. Seipin monomers assemble into a decameric cage-like structure and sit in the ER to provide a space that permits triacylglycerol molecules to interact with each other, rather than with phospholipid acyl chains, a process that is probably aided by trans-membrane protein segments. This enables phase separation of the triacylglycerols, lens formation, and growth to a point where the seipin oligomer opens toward the cytoplasm so the lens can form a budding lipid droplet.
Figure \(\PageIndex{12}\) presents a cartoon that shows the role of ER membrane seipins and other proteins in the assembly of lipid droplets.

In panel (A), seipin is enriched at the endoplasmic reticulum (ER)/lipid droplet (LD) contact sites and is crucial in triglycerides (TAG) flow from the ER to the LD. Seipin interacts with Perilipin1 (PLIN1) and with several TAG synthetic enzymes such as glycerol-3-phosphate acyltransferase (GPAT3), 1-acyl-sn-glycerol-3-phosphate acyltransferase beta (AGPAT2), and LIPIN. No interaction with diacylglycerol acyltransferases (DGAT) has been formally reported. Magré J et al, Int J Mol Sci. 2022 Jan 11;23(2):740. doi: 10.3390/ijms23020740. Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/
In panel (B), representing the fasting state, seipin is enriched at the ER/mitochondria (Mt) contact sites, also named mitochondria-associated membranes (MAM) and participates in ER/mitochondria calcium (Ca2+) flux and mitochondrial activity. Seipin is near MAM Ca2+ regulators IP3(inositol 1,4,5-trisphosphate) receptor (IP3R), voltage-dependent anion channel (VDAC), and sarco-/ER Ca2+ ATPase 2 (SERCA2) as well as glycerol-3-phosphate (G3P), lysophosphatidic acid (LPA), phosphatidic acid (PA), and diacylglycerol (DAG).
Figure \(\PageIndex{13}\) shows an interactive iCn3D model of the S. cerevisiae seipin Homo 10-mer flexible cage at lipid droplet formation sites (7RSL).
.png?revision=1&size=bestfit&width=366&height=190)
Figure \(\PageIndex{13}\): S. cerevisiae seipin homo 10-mer flexible cage at lipid droplet formation sites (7RSL). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...BjYLiQXVJjBpk8
The N-terminal transmembrane segment (TMS) on each monomer is shown in cyan while the C-terminal TMS is shown in magenta. These form the cage sides and top and anchor the complex in the ER membrane. The side chains of a conserved switch sequence (F232xxGLR) on each monomer are shown in spacefill, CPK colors. The gray ribbons represent the part of the complex that is in the lumen of the ER.
Adjacent monomers in the homo 10-mer cage, even though they are identical in sequences, adopt different orientations for two helices that enable a binary switch in the conformation of the complex. Two adjacent subunits, the A chain in blue and the B chain in orange, are shown, along with their overlap, in Figure \(\PageIndex{14}\).
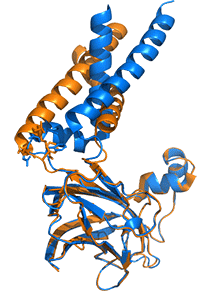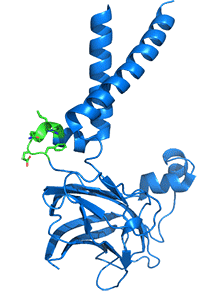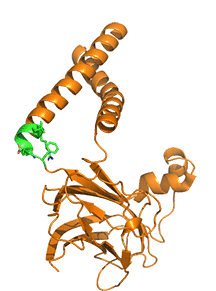
A highly conserved "switch" region is shown in green with its highly conserved motif F232xxGLR shown as sticks with CPK colors. Note the large conformational change in the switch region which occurs with a large conformational change of the two helices. This suggests that there are two overall conformations of the complex, a closed form allowing accumulation of TAGs and an open form which allows the budding and release of lipid droplets. A possible model for the formation and budding of lipid droplets is illustrated in Figure \(\PageIndex{15}\). As in the above figure, the A (open) chain is shown in blue while the B (closed) chain is shown in orange.

Some of the surface proteins on lipid droplets can extend long helical hairpins of hydrophobic peptides deep into the lipid core. For example, perilipins constitute a family of at least five phosphorylated proteins that bind to droplets in animals and share a common region, the so-called ‘PAT’ domain, named for the three original members of the family that include perilipin and ADRP. Proteins related evolutionarily to these are found in more primitive organisms, including insects, slime molds, and fungi, but not in the nematode Caenorhabditis elegans. In mammals, perilipin A (or 'PLIN1' or more accurately the splice variant 'PLIN1a') is a well-established regulator of lipolysis in adipocytes, and it is believed to be involved in the formation of the large lipid droplets in white adipose tissue. The perilipins PLIN1 and PLIN2 have functions in triacylglycerol metabolism in tissues other than adipocytes, and PLIN2 is the main perilipin in hepatocytes; PLIN5 operates in tissues that oxidize fatty acids such as the heart. Other surface proteins of lipid droplets are enzymes intimately involved in triacylglycerol metabolism, although there is a suggestion that cytoplasmic droplets may act as a storage organelle for hydrophobic proteins whose function is elsewhere in the cell.
Lipolysis
When fatty acids are required by other tissues for energy or other purposes, they are released from the triacylglycerols by the sequential actions of three cytosolic enzymes at neutral pH, i.e., adipose triacylglycerol lipase (ATGL), hormone-sensitive lipase (HSL) and monoacylglycerol lipase, which cycle between the cytoplasmic surfaces of the endoplasmic reticulum and the surface layer of lipid droplets. Simplistically, ATGL hydrolyses triacylglycerols to diacylglycerols, which are hydrolyzed by HSL to monoacylglycerols before these are hydrolyzed by the monoacylglycerol lipase to complete the process. Lipolysis proceeds in a highly ordered manner with stimulation through cell-surface receptors via neurotransmitters, hormones, and autocrine/paracrine factors that activate various intracellular signaling pathways and increase kinase activity. A protein perilipin (PLIN1) has been described as "the gatekeeper of the adipocyte lipid storehouse" that regulates lipolysis by acting as a barrier to lipolysis in non-stimulated cells. However, on β-adrenergic stimulation during fasting, it is phosphorylated by the cAMP-protein kinase, which changes its shape and reduces its hydrophobicity, and in the process activates lipolysis. An isoform, perilipin A, is the main regulatory factor in white adipose tissue. However, many other proteins interact with the three enzymes to modulate their activity, location, and stability.
The adipose triacylglycerol lipase, which initiates the process, was discovered surprisingly recently. It is structurally related to the plant acyl-hydrolases in that it has a patatin-like domain in the NH2-terminal region (patatin is a non-specific acyl-hydrolase in potatoes). Specific transport mechanisms guide ATGL from the endoplasmic reticulum membrane to lipid droplets, where it is located on the surface both in the basal and activated states. This lipase is specific for triacylglycerols containing long-chain fatty acids, preferentially cleaving ester bonds in the sn-1 or sn-2 position (but not sn-3), and it yields diacylglycerols and free fatty acids as the main products, with low activity only towards diacylglycerols, and none to monoacylglycerols and cholesterol esters. However, it also has transacylase and phospholipase activities, and it hydrolyses retinol esters in hepatic stellate cells. Adipose triacylglycerol lipase can be activated at the same time as hormone-sensitive lipase and is now believed to be rate-limiting for the first step in triacylglycerol hydrolysis. Figure \(\PageIndex{16}\) shows the sequential hydrolysis of TAGs in adipocytes and lipid droplets.

Regulation of the enzymatic activity is a complex process, and for example, a lipid droplet protein designated Gene identification-58 (CGI-58 or ABHD5), is known to be an important activating factor and is required for hydrolysis of fatty acids from position sn-1. In the resting state, this protein binds to perilipin (PLIN1), but on hormonal stimulation, the latter is phosphorylated leading to dissociation and interaction of CGI-58 with phosphorylated ATGL to commence the first step in triacylglycerol hydrolysis. Mutations in adipose triacylglycerol lipase or CGI-58 are believed to be responsible for a syndrome in humans known as ‘neutral lipid storage disease’. A second protein (G0S2) inhibits the enzyme.
Hormone-sensitive lipase in various isoforms is a structurally unique member of the large Ser-lipase/esterase family of enzymes in animals. It is regulated by the action of the hormones insulin and noradrenaline by a mechanism that ultimately involves phosphorylation of the enzyme by cAMP-protein kinase (as with perilipin), thereby increasing its activity and causing it to translocate from the cytosol to the lipid droplet to initiate the second step in hydrolysis. Its activity is regulated further by a variety of proteins that include PLINs and fatty acid binding proteins (FABP). Hormone-sensitive lipase has a broad substrate specificity compared to other neutral lipases, and in addition to its activity towards triacylglycerols, it will rapidly hydrolyze diacylglycerols, monoacylglycerols, retinol esters, and cholesterol esters. Diacylglycerols are hydrolyzed ten times as rapidly as triacylglycerols. Within the triacylglycerol molecule, hormone-sensitive lipase preferentially hydrolyses ester bonds in the sn-1 and sn-3 positions, leaving free acids and 2‑monoacylglycerols as the main end products.
The monoacylglycerol lipase is believed to be the rate-limiting enzyme in lipolysis, i.e., the final step in triacylglycerol catabolism releasing free glycerol and fatty acids, and it is found in the cytoplasm, the plasma membrane, and in lipid droplets. It is specific for monoacylglycerols, but with no positional specificity, and has no activity against di- or triacylglycerols. As it is the enzyme mainly responsible for the deactivation of the endocannabinoid 2-arachidonylglycerol and is highly active in malignant cancers, it is attracting pharmaceutical interest. A further enzyme, α/β hydrolase containing-6 (ABHD6), is located on the inner leaflet of the plasma membrane and preferentially hydrolyses fatty acids at the sn-1 position over the sn-2 position of monoacylglycerols, and it also hydrolyses lysophospholipids and bis(monoacylglycerol)-phosphate. This has been associated with the development of insulin resistance and the progression of cancer. Additional lipolytic enzymes, including carboxyesterases, are believed to operate against triacylglycerols in cytoplasmic lipid droplets in the liver.
Unesterified fatty acids released by the combined action of these three lipases are exported into the plasma for transport to other tissues in the form of albumin complexes, while the glycerol released is transported to the liver for metabolism by either glycolysis or gluconeogenesis.
Eventually, the whole organelle can disappear, including the proteins, when they undergo a process of autophagy (lipophagy), i.e., the delivery of the organelles to lytic compartments for degradation. This can occur through direct lysosomal invagination or more often by a multistep process involving the formation of double-membrane vesicles termed 'autophagosomes' around droplets with subsequent lysosomal fusion and degradation of the triacylglycerols by the lysosomal acid lipase. This process is important for the regulation of cellular lipid levels in various tissues and disease conditions, especially during starvation. It is relevant to tumorigenesis and cancer metastasis, and to neurodegenerative and neuroinflammatory diseases. While lipophagy is mechanistically distinct from lipolysis, there is cross-talk between the two.
Lipases carry out hydrolysis reactions (nucleophilic substitution with water as the nucleophile), alcoholysis reactions (with an ROH as the nucleophile), and even transesterification reactions with other fatty acid esters. Lipases are critical enzymes yet there are no good structures available for hormone-sensitive lipase for example. Yet much is known about the structure and activity of lipases. Lipases have a catalytic Ser-His-Asp triad like serine proteases, which also carry out hydrolysis reactions. However, the substrates for lipase are nonpolar and could be part of larger structures such as lipid droplets. As such, the lipases work at the interface between aqueous and non-aqueous interfaces. They are interfacially activated.
Lipases appear to have low activity in aqueous solutions but more in solutions that are more nonpolar. Many appear to have two conformations, a closed one with lower activity and poor lipid substrate binding properties, and an open one with higher activity and activation by nonpolar solvents. An alpha-helical lid appears to close off the active site in the closed conformation. In a more nonpolar solvent or the presents of bulk lipids, such as lipid droplets, the alpha-helical lid moves exposing the enzyme's hydrophobic binding site to substrate interactions.
Figure \(\PageIndex{17}\) shows an interactive iCn3D model comparing the structure of a Thermomyces lanuginosus lipase in its closed (1DT3) and its open conformation (1EIN).
_and_is_open_conformation_(1EIN).png?revision=1&size=bestfit&width=232&height=174)
Figure \(\PageIndex{17}\): The structure of a Thermomyces lanuginosus lipase in its closed (1DT3) and its open conformation (1EIN). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/icn3d/share.html?C9yEwy6SR6MZ9Q1S7
The closed structure (1DT3) is shown in magenta and the open (1EIN) is in cyan. The active site triad Ser146, His258, and Asp201 is shown in CPK-colored sticks and labeled for each. The lid is shown in yellow with its side chains as sticks. Toggle between the two states using the "a" key. Note how the lid covers the active site in the closed (1DT3) structure. You will have to rotate the molecule to see the optimal orientation. Also, close out menu boxes for a larger and more centered display.
Endocrine functions
Not only does the adipocyte provide a store of energy but it is an endocrine organ that manages the flow of energy through the formation of the hormone leptin, which signals through the adiponectin receptors (AdipoR)1 and AdipoR2 and binds to the non-signaling interacting protein, T‑cadherin. The result is the stimulation of signaling cascades to communicate with other tissues through the secretion of cytokines and other mediators. The synthesis of leptin is tightly controlled by adipocytes mainly, although it is also produced by the stomach, placenta, and mammary gland, and its principal function is the provision of information on the state of fat stores to other tissues. In so doing, it regulates food intake and energy expenditure. Leptin was initially described as an anti-obesity hormone, but it serves as an adiposity signal with a vital function in maintaining adipose tissue mass to ensure survival under conditions of negative energy balance and so protect against either a deficit or an excess of adiposity. Lipid droplets have a role in this process, sinc perilipin is required for the sensing function.
Adiponectin is a potent insulin sensitizer and suppressor of cell death and inflammation, directly promoting anti-diabetic and anti-atherosclerotic outcomes. Insulin is the main hormone that affects metabolism and its receptor at the plasma membrane is located in caveolae. The release of proinflammatory cytokines can stimulate lipolysis and cause insulin resistance, in turn leading to dysfunction of adipose tissue and systemic disruption of metabolism. Thus, adipose tissue metabolism has profound effects on whole-body metabolism, and defects in these processes can have severe implications for such serious pathological conditions as diabetes, obesity, cardiovascular disease, fatty liver disease, and cancer in humans. It is hoped that the development of specific inhibitors for hormone-sensitive lipase will improve the treatment of such metabolic complications. As caveolae, which contain the proteins caveolins (and presumably sphingolipids) and are particularly abundant in adipocytes, modulate the flux of fatty acids across the plasma membrane and are involved in signal transduction and membrane trafficking pathways, it is evident that they have a major role in this aspect of lipid metabolism.
White fat acts as an endocrine organ and can release a variety of hormones including adipokines (analogous to cytokines, chemokines, lymphokines) adipsin, and leptin as well as tumor necrosis factor α (TNF-α), adiponectin, resistin, and RBP4.
Functions other than energy management
Lipid droplets accumulate within many cell types other than adipocytes, including leukocytes, epithelial cells, hepatocytes, and even astrocytes, especially during infections, cancer, and other inflammatory conditions. They are important for the cellular storage and release of hydrophobic vitamins, signaling precursors, and other lipids that are not related to energy homeostasis while reducing the dangers of lipotoxicity. On the other hand, excessive fatty acid accumulation is associated with lipotoxicity, endoplasmic reticulum stress, and mitochondrial damage and dysfunction, so lipid storage in lipid droplets must be balanced for health.
A variety of enzymes are associated with lipid droplets, including protein kinases, which are involved in many different aspects of lipid metabolism, such as cell signaling, membrane trafficking, and control of the production of inflammatory mediators like the eicosanoids. Lipolysis enables the secretion of lipid species termed lipokines (more generally 'adipokines') from adipocytes that may signal in a hormone-like fashion to other tissues, thereby modulating gene expression and physiological function, including food intake, insulin sensitivity, insulin secretion, and related processes. These include palmitoleic acid (9-16:1) and fatty acid esters of hydroxy fatty acids (FAHFA), though the circulating proteins adiponectin and leptin have been studied more intensively. Adiponectin is a powerful insulin sensitizer and suppressor of apoptosis and inflammation with anti-diabetic and anti-atherosclerotic functions, often operating through its effects on sphingolipids, while leptin exerts most of its effects on the brain to trigger behavioral, metabolic, and endocrine responses to control the body's fuel reserves.
Indeed, there are now suggestions that lipid droplets in all cell types are essential for the response mechanisms to cellular stress, including autophagy, inflammation, and immunity, and act as hubs to integrate metabolic and inflammatory processes. Via their lipolytic machinery, they regulate the availability of fatty acids for the activation of signaling pathways and the production of oxylipins from polyunsaturated fatty acids. For example, triacylglycerols in cytoplasmic lipid droplets of human mast cells, which are potent mediators of immune reactions and influence many inflammatory diseases, have a high content of arachidonic acid and this can be released by adipose triacylglycerol lipases as a substrate for production of specific eicosanoids when the cells are stimulated appropriately. Active metabolism in lipid droplets is important for the differentiation of monocytes, and it is essential for the sustained functional activity of differentiated macrophages, especially in relation to inflammation.
During apoptosis, triacylglycerols enriched in polyunsaturated fatty acids accumulate in lipid droplets, possibly as a protective mechanism against membrane damage caused by oxidative stress and hydroperoxide formation in this process. Triacylglycerols in lipid droplets of the skin are a highly specific source of linoleic acid that is required for the formation of the O-acylceramides, which are essential for epidermal barrier function. An organelle termed the midbody in dividing cells in humans and rodents contains a unique triacylglycerol that is a single molecular species consisting of three fatty acids 16:1-12:0-18:1 (12:0 especially is rarely detected in human lipids), but its function is not known.
Vitamin E (tocopherols) and vitamin A in the form of retinyl esters are stored in cytoplasmic lipid droplets, and the latter are present in appreciable concentrations in the stellate cells of the liver, for example. In endocrine cells of the gonads and adrenals, cholesterol esters stored in lipid droplets are an important source of cholesterol for the mitochondrial biosynthesis of various steroid hormones. In the nucleus of the cell, in addition to providing a reservoir of fatty acids for membrane remodeling, lipid droplets can sequester transcription factors and chromatin components and generate the lipid ligands for certain nuclear receptors.
In addition to their role in lipid biochemistry, lipid droplets participate in protein degradation and glycosylation. Their metabolism can be manipulated by pathogenic viruses and bacteria such as Mycobacterium tuberculosis with unfortunate consequences for the host, but they also serve as reservoirs for proteins that fight intracellular pathogens. In consequence, such lipid droplets and their enzyme systems may be markers for disease states and are also considered to targets for pharmaceutical intervention.
Insects
In insects, the fat body is a multifunctional tissue that is the main metabolic organ. It integrates signals that control the immune system, molting, metamorphosis, and synthesis of hormones that regulate innumerable aspects of metabolism. In fat body cells, lipids, carbohydrates, and proteins are the substrates and products of many pathways for use in energy production or to act as reserves for mobilization at the appropriate stage of life (diapause, metamorphosis, and flight). In relation to innate and acquired humoral immunity, the fat body produces bactericidal proteins and polypeptides, i.e., lysozyme. It is also important in the early stages of an insect's life due to the production of vitellogenin, the yolk protein needed for the development of oocytes.
Brown Adipose Tissue
Most adipose tissue depots ('white fat') serve primarily as storage and endocrine organs that provide a reservoir of nutrients for release when the food supply is low. However, a second specialized form of adipose tissue, brown fat, is multilocular, highly vascularized, and rich in mitochondria and the iron-containing pigments that transport oxygen and give the tissue its color and name. Brown adipocytes arise from progenitor cells that are closer to those of skeletal muscle than white adipocytes. In humans, these depots tend to be located in specific anatomical regions such as subcutaneous areas around the neck, where their function may be to supply warm venous blood directly to the spinal cord and brain, and elsewhere to the heart, kidney, pancreas, and liver. Brown fat can oxidize fat so rapidly that heat is generated (“non-shivering thermogenesis”), and it is especially important in young animals and those recovering from hibernation.
In brief, during cold exposure, the release of noradrenaline and stimulation of β-adrenergic receptors in the nervous system initiates a catabolic program that commences with a rapid breakdown of cellular triacylglycerol stores and release of unesterified fatty acids and transient activation of a co-activator of peroxisome proliferator-activated receptor gamma (PPARγ). These set in motion a signaling process that results in the efficient β-oxidation of fatty acids to produce heat. The key molecule is believed to be the uncoupling protein-1 (UCP1), which acts as a valve to uncouple electron transport in the respiratory chain from ATP production with a highly exothermic release of chemical energy, i.e., as heat rather than as ATP. Its actions are illustrated in Figure \(\PageIndex{18}\).
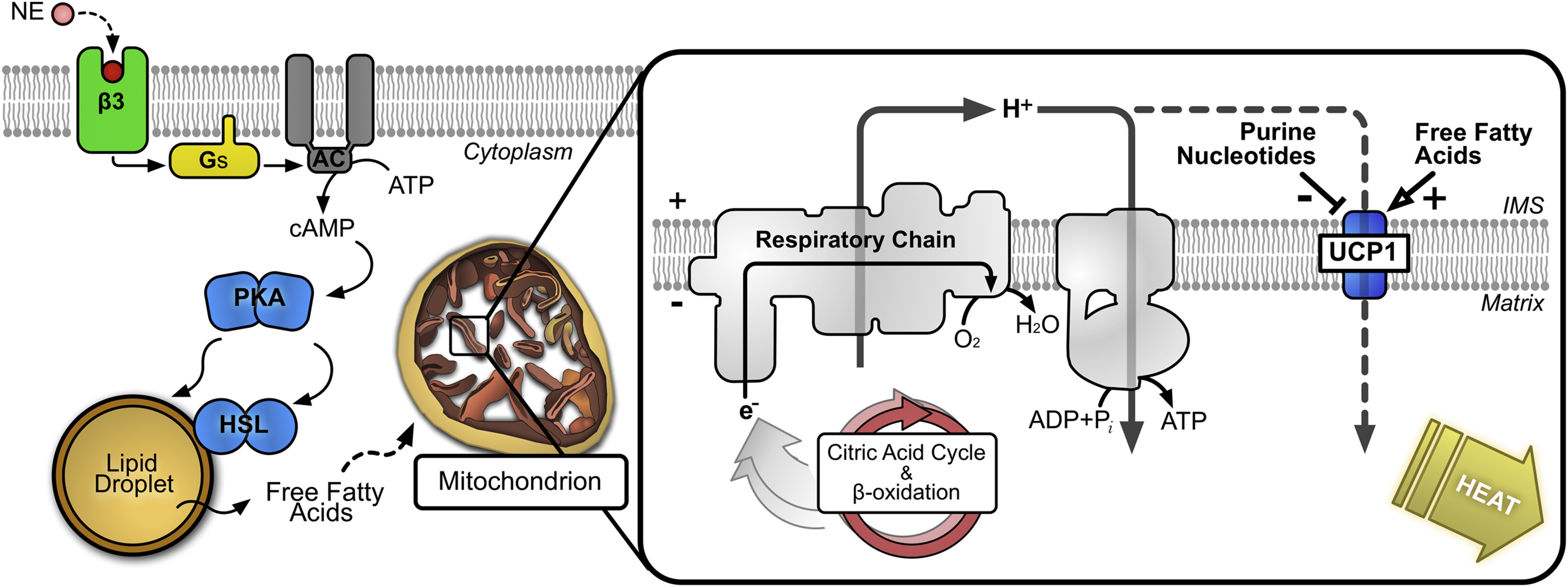
Figure \(\PageIndex{18}\): Role of UCP1 in generation of heat. https://commons.wikimedia.org/wiki/F...n_the_cell.jpg. Creative Commons Attribution-Share Alike 4.0 Internationa
This is an example of a futile cycle that releases heat. Although many aspects of the mechanism are uncertain, it is clear that proton conductance by UCP1 is highly regulated and inducible. It is activated by free long-chain fatty acids and inhibited by purine nucleotides, i.e., fatty acids are not only the substrate for thermogenesis but act also as self-regulating second messengers. The mitochondrial phospholipid cardiolipin, which is intimately involved in oxidative phosphorylation, is indispensable for stimulating and sustaining the function of thermogenic fat. Upon activation of brown adipose tissue, the dense vasculature increases the delivery of fatty acids and glucose to the brown adipocytes and warms the blood passing through the tissue.
Long-chain fatty acids bind to UCP1 and may be transported through the membrane along with a proton. Alternatively, it may just bind and activate the transport of a proton. Figure \(\PageIndex{19}\) shows an interactive iCn3D model of the AlphaFold predicted structure of human UCP1 (P25874).
.png?revision=1&size=bestfit&width=304&height=397)
Figure \(\PageIndex{19}\): AlphaFold predicted structure of human UCP1 (P25874). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...FkfrEqWE3D2dX6
Imagine a bilayer orienting perpendicularly to the gray helices representing the transmembrane helical segments that imbed the protein into the inner mitochondrial membrane. The spacefill Ile150 denotes the matrix domain. Two key residues (K56, K29) involved in the binding of the carboxyl end of the fatty acids are shown on the matrix side of the protein is CPK-colored sticks and labeled. The black faces of the helices in the inner membrane region show the most probable binding site for the acyl chain of the fatty acid. The lighter yellow/orange regions have greater predicted uncertainty in the AlphaFold computational model.
There is evidence that acylcarnitines produced in the liver from fatty acids released from white adipose tissue in response to cold exposure are transported in plasma to brown adipose tissue and can serve as a substrate for thermogenesis. Indeed, a wide array of circulating lipids contributes to thermogenic potential, including free fatty acids and triacylglycerols.
The activities of acyl-CoA synthetases and acyl-CoA thioesterases determine the availability of substrates for β-oxidation and consequently the thermogenic capacity. Synthesis of the lipokine (or 'batokine') octadecanoid 12,13-dihydroxy-9Z-octadecenoic acid (12,13-diHOME), is induced by cold also, and this stimulates the activity of brown adipose tissue by promoting the uptake of fatty acids, acting via G-protein-coupled receptors. It increases cardiac function and cardiomyocyte respiration via enhanced calcium cycling. Similarly, there are suggestions that n‑3 polyunsaturated fatty acids may promote adaptive thermogenesis, for example through the activity of the 12‑lipoxygenase metabolite and batokine 12‑hydroxyeicosapentaenoic acid (12-HEPE) by improving glucose metabolism via increased glucose uptake into adipocytes and skeletal muscle. (FAHFA) are relevant in this context. Peroxisomal synthesis of plasmenyl-phospholipids is believed to regulate adipose tissue thermogenesis by mediating mitochondrial fission.
In hibernating mammals, brown adipose tissue is especially important metabolically, and even in laboratory animals such as mice, it can consume about 50% of dietary lipids and glucose when the animals are exposed to cold. Similarly, in even humans, the activity of brown adipose tissue is induced acutely by cold and is stimulated via the sympathetic nervous system, and the relevance of this tissue to human metabolism is now becoming apparent. For example, there are suggestions that brown adipose tissue can behave as an endocrine system to secrete endocrine factors ('batokines') that may be favorable in reducing cardiovascular risk. For obvious reasons, there are efforts to determine whether sustained activation of brown fat by pharmaceutical means could be beneficial for human disease states, including obesity, diabetes, and cardiovascular disease.
Such research has been stimulated by the observation that clusters of distinct adipocytes with thermogenic capacity in addition to their storage function can be present in white adipose tissue and emerge in response to various physiological signals, especially reactive oxygen species. They are termed 'beige or brite' adipocytes and arise from multipotent pre-adipocytes. Brown adipose tissue is a discrete organ in animals, but beige adipose tissue is interspersed with white, and the two forms have different developmental origins. In adult humans, most of the deposits once thought to be classical brown adipose tissue do not contain the genetic markers for this tissue and are now recognized to be beige/brite fat, although brown adipose tissue per se is present in significant amounts in human newborns and infants. However, beige adipocytes utilize the same machinery to release heat by oxidation of fatty acids under β-adrenergic stimulation.
The uncoupler protein 1 (UCP1) appears to be expressed only in brown and beige fat cells and is a marker for those cells. It can be regulated transcriptionally and posttranscriptionally by ROS modification of a cysteine to produce a sulfenyl group. The more classical brown adipocyte arises developmentally from the skeletal muscle-like cell, whereas the beige appears to develop to some extent from vascular smooth muscle cells. The beige cells can differentiate into more white adipocytes or more brown ones, by the factors described in Figure \(\PageIndex{20}\).

Figure (\PageIndex{20}\): Factors altering beige adipose tissue. Paul Cohen and Bruce M. Spiegelman. Molecular Biology of the Cell, 27, 2017. https://doi.org/10.1091/mbc.e15-10-0749. Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
In addition to the type of fat cell, the location and size of the adipocyte also affect health risks. Obesity, cardiovascular disease, type II diabetes, and nonalcoholic fatty liver disease all depend on and are correlated with fat cell dysfunction. The endocrine role of adipose tissue plays a key role in the development of dysfunction. The main depots of fat are the subcutaneous adipose tissue (SAT) and the visceral adipose tissue (VAT), the latter of which is associated with proinflammatory effects and associated greater health risks (type II diabetes, hypertension, metabolic syndrome, obesity, etc.). Figure (\PageIndex{21}\) below summarizes the health consequence as the size of the SAT and VAT adipocytes increase (in the direction of the red arrow).

Figure (\PageIndex{2}\): Adipocyte characteristics and cardiovascular risk. The diagram describes a proposed model, based on the results of adipocyte size/source, potential interactions with metabolic mediators, and pathophysiological effects. SAT, subcutaneous adipose tissue; VAT, visceral adipose tissue; HbA1c, glycated hemoglobin (a marker of elevated blood sugar associated with diabetes); RES, resistin; ADIPON, adiponectin; INS, insulin; HDLc; high-density lipoprotein cholesterol; FMD, flow-mediated dilation; CIMT, carotid intima media thickness. Suárez-Cuenca et al. Sci Rep 11, 1831 (2021). https://doi.org/10.1038/s41598-021-81289-2. Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/.
It appears that the larger the adipocyte, the greater the inflammatory response. This condition is called adipocyte hypertrophy and it may be more important than obesity per se as a risk factor for inflammation, as indicated by inflammatory cytokine levels and macrophage infiltration into adipose tissue. The adipose-derived hormone leptin, when released under normal conditions, leads to appetite suppression and also decreases liver and muscle fat. Higher fat storage is associated with increased blood leptin levels. It also correlates with greater lymphocyte size.
In contrast, the adipose hormone adiponectin leads to positive health consequences including increased sensitivity to insulin, decreased inflammation, and the formation of adipocytes from stem cells (adipogenesis). The larger the fat cell, the lower the serum levels of adiponectin. However, the link between fat cell size and adiponectin levels may be determined more by obesity.
Other Functions of Triacylglycerol Depots
Subcutaneous depots act as a cushion around joints and serve as insulation against the cold in many terrestrial animals, as is obvious in the pig, which is surrounded by a layer of fat, and it is especially true for marine mammals such as seals. Those adipocytes embedded in the skin differ from the general subcutaneous depots and support the growth of hair follicles and regenerating skin, and they may also have a defensive role both as a physical barrier and by responding metabolically to bacterial infection.
In marine mammals and fish, the fat depots are less dense than water and so aid buoyancy with the result that less energy is expended in swimming. More surprisingly perhaps, triacylglycerols together with the structurally related glyceryl ether diesters and wax esters are the main components of the sonar lens used in echo location by dolphins and toothed whales. The triacylglycerols are distinctive in that they contain two molecules of 3‑methylbutyric (isovaleric) acid with one long-chain fatty acid. It appears that the relative concentrations of the various lipids in an organ in the head of the animals (termed the ‘melon’) are arranged anatomically in a three-dimensional topographical pattern to enable them to focus sound waves.
In cold climates, many insects do not feed over winter and must manage their energy stores to meet the energetic demands of development and reproduction in the spring. Some insect species that are tolerant of freezing produce triacylglycerols containing acetic acid, and these remain liquid at low temperatures; by interacting with water, they may play a role in cryoprotection.
Triacylglycerol Metabolism in Plants and Yeasts
Fruit and seed oils are major agricultural products with appreciable economic and nutritional value to humans. The mesocarp of fruits is a highly nutritious energy source that attracts animals that help to disperse the seeds, and in plants such as the oil palm and olive trees a high proportion of the fruit flesh contains triacylglycerols. Similarly in seeds, triacylglycerols are the main storage lipid and can comprise as much as 60% of their weight. Fruit lipids are not intended for use by the plant per se and are stored in lipid droplets in large irregular structures that break down readily, but seed lipids are required for the development of the plant embryo, so their metabolism is of particular importance. However, triacylglycerol biosynthesis and metabolism are required also for pollen viability and to maintain lipid homeostasis in chloroplasts (see the note on plastoglobules below).
Seed oils: In seeds and other plant tissues, biosynthesis of fatty acids takes place in plastids, and these are stored in the form of triacylglycerols in lipid droplets with a coherent surface layer of proteins and lipids in the embryo (e.g., Arabidopsis, soybean or sunflower) or endosperm (e.g., castor bean) tissues of seeds. In addition to the common range of fatty acids synthesized in plastids, mainly palmitate, and oleate, some plant species produce novel fatty acids, including medium- and very-long-chain components and those with oxygenated and other functional moieties. A specific means of diverting these to seeds for triacylglycerol production exists to prevent disruption of the plant membranes. Seed development occurs in three stages - rapid cell division with no accumulation of storage material, rapid deposition of triacylglycerols and other energy-rich metabolites, and finally desiccation.
During the period of oil accumulation in seeds, the newly formed ACP esters of fatty acids are first hydrolyzed by two different classes of acyl-ACP thioesterases at the inner plastid envelope membrane, before the unesterified fatty acids are transported to the endoplasmic reticulum (ER), by a family of fatty acid export proteins (FAX) of which there are seven isoforms in Arabidopsis, two of which (FAX2 and FAX4) are highly expressed during the early stage of seed development. The unesterified fatty acids are shuttled across the plastid outer envelope, probably by vectorial acylation by long-chain acyl-CoA synthases, which catalyze the formation of CoA esters.
In the ER, triacylglycerols and membrane lipids are synthesized by the Kennedy and other pathways described above. In yeast and plants, 1,2‑diacyl-sn-glycerol esterification is the only committed step in triacylglycerol production, and this occurs by mechanisms that can be both dependent or independent of acyl-CoA esters. The acyl-CoA-dependent route is catalyzed by diacylglycerol:acyl-CoA acyltransferases (DGATs) with acyl-CoA and diacylglycerols as substrates, and two membrane-bound isoenzymes (DGAT1 and DGAT2) and a cytosolic isoenzyme (DGAT3) are known, DGAT1 is a key enzyme involved in triacylglycerol formation in developing seeds, while DGAT2 is especially important in those plant species with unusual fatty acid compositions. In Arabidopsis, DGAT3 has some specificity for polyunsaturated fatty acids in seed development. Figure \(\PageIndex{22}\) summarizes TAG synthesis in plant ER.

In addition, a substantial proportion of triacylglycerol biosynthesis in some plant species is synthesized by flux through the membrane phospholipid phosphatidylcholine, produced by what are sometimes termed inaccurately the 'eukaryotic and prokaryotic pathways' with differing positional distributions, in which diacylglycerols are generated from phosphatidic acid by the action of a phosphatidate phosphatase as an intermediate. In the acyl-CoA-independent reaction, the direct transfer of one fatty acid from phosphatidylcholine to diacylglycerol by the action of the phospholipid:1,2-diacyl-sn-glycerol-acyltransferase (PDAT) enzyme also occurs, with the formation of lysophosphatidylcholine as a byproduct, which can be re-esterified for further reaction. As phosphatidylcholine undergoes extensive remodeling and its fatty acid components are subject to modification, for example by desaturation to form linoleic and linolenic acids, the compositions and especially the positional distributions of triacylglycerols produced in this way can be very different from those synthesized by the ‘classical’ pathways. Phosphatidylcholine may also function as a carrier for the trafficking of acyl groups between organelles and membrane subdomains, and it has been suggested that an assembly of interacting enzymes may facilitate the transfer of polyunsaturated fatty acids from this phospholipid to triacylglycerols in seeds. Both sterol and sphingolipid biosynthesis appear to be important factors for efficient seed oil production.
As triacylglycerol synthesis continues, oil droplets accumulate between the leaflets of the endoplasmic reticulum and are surrounded by a monolayer of phospholipids, sterols, and proteins, which in Arabidopsis include oleosins, a caleosin, a steroleosin, a putative aquaporin, and a glycosylphosphatidylinositol-anchored protein. Oleosins are the most abundant of these (~65%) and are small proteins (15-30 kDa) that contain cytosolic-facing N- and C-termini and a large hydrophobic domain necessary to target them to lipid droplets, where they are important for control of their size and stability. Eventually, lipid droplets “bud off” from the endoplasmic reticulum with their monolayer of phospholipids and proteins, and they are released into the cytosol by a yet-to-be-defined mechanism.
At the onset of germination, water is absorbed, and esterases/lipases are activated. The process of lipolysis begins at the surface of oil bodies, where the oleosins, which are the most abundant structural proteins, are believed to assist in the docking of lipases and to control the size and stability of lipid droplets in seeds. A number of esterases/lipases have been cloned from various plant species and possess a conserved catalytic triad of Ser, His, and Asp or Glu, somewhat different from the animal lipases, as in patatin (an especially abundant lipolytic protein in potatoes), which can hydrolyze triacylglycerols but not phospho- or galactolipids. The most important of these is believed to be the 'sugar-dependent lipase 1 (SDP1)', which is a patatin-like lipase similar in function to the mammalian adipose triacylglycerol lipase discussed above and is located on the surface of the oil body. This is active mainly against triacylglycerols to generate diacylglycerols but presumably works in conjunction with di- and monoacylglycerol lipases to generate free fatty acids and glycerol.
The lipid droplets in seeds exist near glyoxysomes (broadly equivalent to peroxisomes). These are the membrane-bound organelles that contain most of the enzymes required to oxidize fatty acids derived from the triacylglycerols via acetyl-CoA to four-carbon compounds, such as succinate, which are then converted to soluble sugars to provide germinating seeds with energy to fuel the growth of the seedlings and to produce shoots and leaves. In addition, they supply structural elements before the seedlings develop the capacity to photosynthesize. How the products of lipolysis are transported to the glyoxysomes for further metabolism has still to be determined, but a specific ‘ABC’ transporter is required to import fatty acids into the glyoxysomes in Arabidopsis. The free acids are converted to their coenzyme A esters by two long-chain acyl-CoA synthetases located on the inner face of the peroxisome membrane before entry into the β‑oxidation pathway. All these processes are controlled by an intricate regulatory network, involving transcription factors that crosstalk with signaling events from the seed maturation phase through to embryo development. After about two days of the germination process, the glyoxysomes begin to break down, but β-oxidation can continue in peroxisomes in leaf tissue.
Lipid droplets - plastoglobules: Triacylglycerol-rich lipid droplets (LD) have been observed in most cell types in vegetative tissues of plants as well as in seeds, and although their origin and function are poorly understood, they contain all the enzymes required for triacylglycerol metabolism together with phospholipases, lipoxygenases, and other oxidative enzymes. Instead of oleosins, these lipid droplets in plants and algae contain a family of ubiquitously expressed 'LD-associated proteins' on the surface, together with a monolayer of phospholipids (mainly phosphatidylcholine), galactolipids such as sulfoquinovosyldiacylglycerol and in some species betaine lipids. As in yeast and humans, seipins (three in Arabidopsis) are necessary for normal LD biogenesis. Again, LD forms within the bilayer of the endoplasmic reticulum and pinch off into the cytoplasm. Abiotic stresses can induce the remodeling of lipid membranes through lipase action with the formation of toxic lipid intermediates, and these can be sequestered by triacylglycerols in lipid droplets to inhibit membrane damage and potentially prevent cell death. While they are believed to be involved mainly in stress responses, lipid droplets may have other specialized roles, for example in anther and pollen development, where triacylglycerols serve as a source of fatty acids for membrane biosynthesis. Fatty acids derived from triacylglycerols in lipid droplets are believed to be subjected to peroxisomal β‑oxidation to produce the ATP required for stomatal opening and no doubt many other purposes.
In addition, lipid droplets that have been termed 'plastoglobules' are produced by a localized accumulation of triacylglycerols and other neutral lipids between the membrane leaflets of the thylakoid cisternae and then pinch off into the stroma, where they are involved in a wide range of biological functions from biogenesis to senescence via the recruitment of specific proteins. During senescence, for example, lipid droplets accumulate rapidly in the leaves of A. thaliana. In reproductive tissues, may have a more direct function by recruiting and transporting proteins, both for organ formation and successful pollination. Antifungal compounds such as 2-hydroxy-octadecatrienoic acid and other oxylipins are produced from α-linolenic acid in these organelles, and it has been suggested that the latter function as intracellular factories to produce stable metabolites via unstable intermediates by concentrating the enzymes and hydrophobic substrates efficiently. Plastoglobules are also implicated in the biosynthesis and metabolism of vitamins E and K.
Microalgae: Triacylglycerol metabolism in lipid droplets in microalgae is under intensive study because of their potential for nutraceutical and biodiesel production. It seems that similar processes occur in higher plants, but with a simpler genome encoding few redundant proteins. In the unicellular green model microalga Chlamydomonas reinhardtii, for example, key lipid droplet proteins, lipases, and enzymes of β-oxidation have been characterized.
Yeasts: Lipid droplets in yeast are a highly dynamic and functionally diverse hub that ensures stress resistance and cell survival by promoting membrane and organelle homeostasis. As most of the important biosynthetic and catabolic enzymes involved in triacylglycerol metabolism are conserved between yeasts and mammals, the former proving to be useful models for the study of triacylglycerol production. The size and triacylglycerol content of lipid droplets in yeasts change appreciably in different stages of growth and development, and Saccharomyces cerevisiae contains a single phosphatidic acid phosphatase (Pah1), which has an essential role in this process. During vegetative growth, Pah1 in the cytosol is phosphorylated by multiple protein kinases, and this enables the synthesis of phospholipids rather than triacylglycerols. As cells progress into stasis, the Pah1 is dephosphorylated and translocates to the endoplasmic reticulum, which ultimately leads to triacylglycerol synthesis for storage in lipid droplets. Some fatty acids derived from phospholipids are utilized for triacylglycerol biosynthesis at the inner nuclear membrane, and this is important for nuclear integrity.
Triacylglycerol Metabolism in Prokaryotes
The study of the biosynthesis of triacylglycerols in bacteria has been stimulated by an awareness of the role of this lipid class in the pathogenesis of Mycobacterium tuberculosis and the relationship with antibiotic biosynthesis by Streptomyces coelicolor. For example, triacylglycerols are believed to be an energy reserve for the long-term survival of M. tuberculosis during the persistence phase of infection as well as a means by which unesterified fatty acids are detoxified. Increasing numbers of bacterial species, for example from the genera Mycobacterium, Nocardia, Rhodococcus, Micromonospora, Dietzia, and Gordonia, are now known to produce triacylglycerols (sometimes wax esters), and these can be stored in lipid droplets in the organisms. The first three steps in triacylglycerol biosynthesis are catalyzed by GPAT, LPAT, and PAP enzymes comparable to those in other organisms. However, it has become apparent that the DGAT can be a dual-function CoA-dependent acyltransferase known as wax ester synthase/diacylglycerol acyltransferase, which accepts a broad diversity of acyl-CoA substrates for esterification of diacylglycerols or long-chain fatty alcohols for the synthesis of triacylglycerols or wax esters, respectively, depending on which intermediates are present in the organisms. Bacteria that lack such an enzyme are unable to produce these non-polar lipids.