
21.1: Biosynthesis of Fatty Acids and Eicosanoids
- Page ID
- 15173


Introduction
We present the full fatty acid synthase pathway based on the structure of the yeast fatty acid synthase (FAS) complex, whose full structure is known. Then we will explore each step in more detail. The mammalian FAS complex is a bit different and we will summarize those differences next. We will end with a summary of the bacterial pathway.
Let's look at the net reaction first and then go back to the structure and mechanism from that. Equation 1 below shows the net reaction.
\[8 \text { Acetyl-CoA }(2 \mathrm{C})+14 \mathrm{NADPH}+14 \mathrm{H}^{+}+7 \mathrm{ATP} \rightarrow 1 \text { Palmitate }(16: 0)+8 \mathrm{CoA}+6 \mathrm{H}_{2} \mathrm{O}+14 \mathrm{NADP}^{+}+7 \mathrm{ADP}+7 \mathrm{Pi}\]
Eight 2C-acetyl-CoAs condense to make the 16:0 fatty acid palmitate. That acetyl-CoA is used to synthesize and elongate fatty acids suggests an immediate explanation for the fact that most fatty acids have an even number of carbon atoms. The net reaction is a reductive biosynthesis making one long molecule from multiple short ones. Hence there must be an energy source to drive the reaction (7ATPs) and a reducing agent (typically NADPH). This reaction takes place in the cytosol and uses the nicotinamide-based redox pair (NADPH/NADP+). These factors differentiate fatty acid synthesis from beta-oxidation, which occurs in the mitochondria and which uses the redox pair NAD+/NADH.
From a chemical perspective, the acetyl-CoAs, high-energy molecules with respect to their hydrolysis products, must be activated using ATP in some way to drive C-C bond formations. That reaction is catalyzed by the enzyme acetyl-CoA carboxylase, which uses the ATP already mentioned and the bound cofactor biotin that adds CO2 to acetyl-CoA to form malonyl-CoA. Neither malonyl-CoA nor CO2 is shown in the above equation since they are present on both sides of the net reaction and cancel out from the final balanced equation. C-C bond formation occurs on the addition of the growing acyl group with the 3C malonyl with the actual addition reaction driven by the release of CO2. The malonyl-CoA produced by acetyl-CoA carboxylase feeds now into the fatty acid synthesis cycle.
Only one other enzyme complex is required for the entire reaction, the fatty acid synthase (FAS) complex. Don't let that fool you into thinking the mechanism is simple though! There are two types of fatty acid synthase complexes, I and II. Type II FAS is found in plants, most bacteria, and in mitochondria. (Its presence in the mitochondria might seem odd since fatty acid oxidation occurs there. We will discuss that in a bit.) There are multiple separate enzymes to catalyze the synthesis of fatty acids in the Type II FAS systems. We won't discuss those. Instead, we will focus on the Type I FAS complex found in some bacteria, fungi like yeast, and higher eukaryotes. The Type I FAS is one very large complex. In mammals, it consists of two α chains with a total molecular weight of 540,000. In yeast, it is an α6β6 heterododecamer with a molecular weight of 2.6 million. In either complex, the alpha and beta chains have multiple different enzyme catalytic domains.
Large enzymatic complexes typically allow bound substrates to proceed to products without dissociation, with the bound intermediates moving through "channels" to the next active site. In the case of Type I FAS, the growing intermediate, which eventually reaches the end product size 16:0 is tethered to an acyl-carrier protein (ACP). The tethered intermediate can "swing" from one active site to another to allow the iterative stepwise chemical reactions to complete before the addition of another 2C acetyl CA to the growing chain. The actual extra 2Cs come from the 3C malonyl-CoA (formed by the extracyclic (outside of the pathway) enzyme acetyl-CoA carboxylase) and immediate release of CO2 which helps drive the reaction forward.
Now we can introduce the fatty acid synthase cycle and feeder acetyl-CoA carboxylase reaction in its entirely, as shown in Figure \(\PageIndex{1}\) for yeast FAS1.

This is a difficult figure to understand but keep in mind that it scales down the complexity of FAS1 enormously. Let's deconstruct the figure to make it more understandable.
The dotted red boxes show the substrates and products that match chemical equation 1. The substrate acetyl-CoA comes into the cycle in two places shown in the extracyclic section to the upper left. It comes in as acetyl-CoA from reaction 3 after attachment to the acyl carrier protein (part of FAS1). It also comes in "effectively" after it has been carboxylated by acetyl-CoA carboxylase in reaction 2 to malonyl-CoA, which enters the cycle as malonyl-ACP after reaction 5. The other substrates/products include CoAs the NAD(P)H/NADP+ couples and H2O shown around the right-hand side of the circular fatty acid synthesis cycle (2 -6 o'clock positions) as well as the final product, 16:0-ACP (9:30 clock position), which forms 16:0-SCoA.
The individual enzymatic domains in the α6β6 hetero-dodecamer yeast FAS1 are shown in colored spheres as shown in Figure \(\PageIndex{2}\).

Figure \(\PageIndex{3}\) summarizes pictorially the repetitive Claisen condensation reactions of acetyl-ACP and malonyl-ACP in the first step, and the growing acyl-ACP chain and malonyl-CoA in the next two cycles.

In each step, the condensation (C-C bond formation) is driven by the decarboxylation of malonyl-CoA. (Of course, earlier expenditure of ATP in step 1 is required). Electrons in malonyl-CoA that form a bond with acetyl-ACP in the first condensation step and acyl-ACP in subsequent condensations are shown as bold blue lines. Note that the condensation is from the head of the malonyl-ACP to the tail of the elongating acyl chain. The ACP or equivalent chains are in the interior of the complex which allows movement of the acyl groups attached to pantetheine chains to interactively reach nearby catalytic domains for each step in the cycle.
Mechanism of individual reactions
Now we can explore some of the reactions in more detail. Then we will look at the structure of yeast FAS and its domain organization.
Reaction 1: Phosphopantetheine transferase
Holo-(acyl carrier protein) synthase (AcpS) from Bacillus subtilis is a member of the phosphopantetheinyl transferase superfamily. AcpS post-translationally modifies ACP to its holo form to activate it. AcpS catalyzes the transfer of the 4'-phosphopantetheinyl (P-pant) moiety of coenzyme A to a serine residue on the ACP. This gives the activated ACP enzyme and adenosine 3'5'-bisphosphate as products. This process is important as ACP enzymes play important roles in several biosynthetic pathways, such as the synthesis of fatty acids, and vitamins, AcpS is essential in the initiation of the biosynthesis of fatty acids, polyketide antibiotics, and non-ribosomal peptide

Figure \(\PageIndex{4}\): https://www.ebi.ac.uk/thornton-srv/m-csa/entry/152/
Reaction 2: Acetyl-CoA Carboxylase (ACC)
This enzyme catalyzes the carboxylation of acetyl-CoA to malonyl-CoA. which then enters the fatty acid synthesis cycle. This is a key enzyme as it is the rate-limiting step, and is regulated. The reaction has two steps:
- carboxylation of biotin on a biotin carboxyl carrier domain of the enzyme at the expense of ATP hydrolysis. The carboxylation, using bicarbonate as a substrate, leads to the formation of a molecule with high energy (with respect to its hydrolysis product), the carboxylated biotin intermediate. (Remember, there is no such thing as a "high energy" bond.)
- transfer of the carboxyl group to acetyl-CoA to form malonyl-CoA which requires the formation of a C-C covalent bond.
Biotin, an essential nutrient and cofactor, "carries" activated carboxyl groups for transfer. It is a carboxyl (not acetyl) donor. It is linked to a lysine side chain in the protein through an amide link.
The enzyme is downregulated by 16:0-SCoA, the end product of the pathway for fatty synthesis by FAS, and by phosphorylation by kinase activation through the cAMP pathways from glucagon binding. You don't want to synthesize fatty acids when your energy state is low (signaled by increases in glucagon). Its activity is upregulated by the binding of citrate to an allosteric site. This makes sense since high citrate, a citric acid cycle intermediate, signifies abundant energy reserves are available so fatty acids would be synthesized for future needs. The dephosphorylated form of the enzyme would signify the need to increase fatty acid synthesis.
Figure \(\PageIndex{5}\) shows a few steps in the carboxylation of biotin and the subsequent transfer of the carboxy group to acetyl-CoA, forming malonyl-CoA.

BCCP represents the biotin carboxyl carrier domain of acetyl-CoA carboxylase.
Biotin carboxylases are found in many enzymes and pathways, not just fatty acid synthesis. Scientists are trying to devise new pathways and enhanced carboxylases to pull CO2 from the air into biosynthetic reactions producing fuels, which could be burned. This would release CO2 back into the atmosphere, in a process that would theoretically but perhaps not practically carbon neutral with respect to greenhouse gas emission.
The abbreviated mechanism shown above shows two of the three activities of acetyl-CoA carboxylase. They are biotin carboxylase (BC) and carboxyltransferase (CT) activity. The third is the biotin carboxyl carrier protein (BCCP) which links biotin to it through a lysine side chain. Different enzymes have different acceptors (acetyl-CoA, pyruvate, etc) of the activated carboxy group so the structure of those centers varies significantly. The BC and BCCP components are structurally similar for different carboxylases.
A mechanism for the E. Coli biotin carboxylase component of the reaction is shown in Figure \(\PageIndex{6}\).

Glu 296 acts as a general acid/base while Arg 338 stabilizes negative charge in intermediates and transition states.
Structure of acetyl-CoA carboxylase (ACC)
There are two forms of ACC, soluble cytosolic ACC1 and mitochondrial membrane-associated ACC2 (the latter which regulates beta-oxidation of fatty acids. The overall domain structure of yeast acetyl-CoA carboxylase is in Figure \(\PageIndex{7}\).

Panel (a) shows the domain organization of ScACC. The domains are labeled and given different colors. The five domains of ACC Central (AC1–AC5) are labeled 1–5. The phosphorylation site in the central region is indicated. The phosphorylation site before the biotin carboxylase (BC) domain core is indicated with the dashed lines, as it is absent in ScACC.
Panel (b) shows the structure of the ScACC holoenzyme dimer [7]. One protomer is shown as ribbons, while the other as a surface. The domains in the monomers are colored according to panel (a) and labeled. Ser1157 (red star) is located in a loop missing in the structure (dashed lines), and its distances to the BC and carboxyltransferase (CT) active sites (black asterisks) and the BC dimer interface (black rectangle) in the holoenzyme are indicated.
A chain domain structure 5CSKA_Yeast Acety-lCoA Carboxylase is shown in Figure \(\PageIndex{8}\).
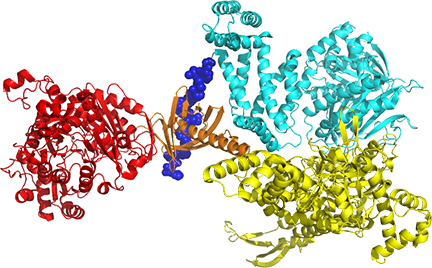
The colors used to show the domains are matched to Figure \(\PageIndex{x}\) above and are as follows:
- Red - BC, biotin carboxylase (has enzymatic activity)
- Gold - BT, an interaction domain
- Blue - BCCP (biotin carboxyl carrier protein (disordered blue spheres)
- Cyan - AC (central bridging noncatalytic domain comprising AC1-AC5).
- Yellow - CT, carboxyl transferase (has enzymatic activity), comprising the N and C subdomains
A similar domain structure is found in human ACC1. The central bridging domain in human ACC1 is called the Central Domain (CD) with four parts, CDN, CDL, CDC1, and CDC2.
Regulation of ACC
ACC1 activity is regulated by product inhibition, allosteric effector, and phosphorylation by different kinases. The enzyme is inhibited by its product, malonyl-CoA, and also the end product of fatty acid synthesis, 16:0-CoA.
The protein is inactive as a monomer and is active as a dimer. Anything that can perturb that equilibrium can affect ACC activity. Phosphorylation generally inhibits the protein by promoting the dissociation of the active dimer. The active dimer, in the presence of the allosteric activator, citrate can aggregate to form fibrils, which are even more active than the dimer.
The protein is post-translationally modified by phosphorylation at several sites but a few stand out. AMP-activated protein kinase (AMPK) phosphorylates human ACC at AMP-activated protein kinase at Ser 80 (in the BC domain). In yeast, the kinase SNF1 (equivalent to human AMPK) does not phosphorylate Ser 80 but it phosphorylates a key Ser 1157 in the AC subdomain AC4 in yeast.
- In humans, phosphorylation at Ser 80 in the BC domain prevents dimerization of the protein, through the BC domain and hence inhibits activity;
- In yeast, phosphorylation st Ser 1157 in AC subdomain AC4 in yeast produces a conformational change in the BC domain region that appears to lead to dimer dissociation.
Human ACC is phosphorylated at a multitude of sites with Ser 80 and 1201 (which is phosphorylated both by AMPK and PKA) being the most important in regulation. In addition, it is phosphorylated at Ser 1263 by a cyclin-dependent protein kinase (CDK). When phosphorylated, the pSer 1263 facilitates the binding of BRAC1, the tumor suppressor protein which when mutated can dramatically increase breast cancer. The binding of BRAC1 leads to an inactive fibril form of ACC. Figure \(\PageIndex{9}\) summarizes the regulation of human CoA carboxylase.

5. Malonyl-Palmitoyl Transferase (MPT)/Malonyl-CoA-acyl carrier protein transacylase (MAT)
These protein activities are found in the yeast (MPT) and human (MAT) domains. The mechanism is shown in Figure \(\PageIndex{10}\).

Two key catalytic residues, Ser 92 and His 201, are involved in an acylation/deacylation of the catalytic serine.
The mechanism is shown in Figure \(\PageIndex{11}\).

Reaction 9. Enoyl Reductase
This is the last reaction in the fatty acid synthase cycle, which of course repeats until a 16:0-SCoA is made. The yeast enzyme is different in that it has a tightly-bound FMN which is involved in electron transfer (reduction) along with NADPH. The mechanism is likely ping-pong with NADP+ released before the enoyl acyl-CoA binds.
Figure \(\PageIndex{12}\) shows an interactive iCn3D model of the enoyl-acyl carrier protein reductase (ER) in complex with NAD+ and triclosan (1QSG)
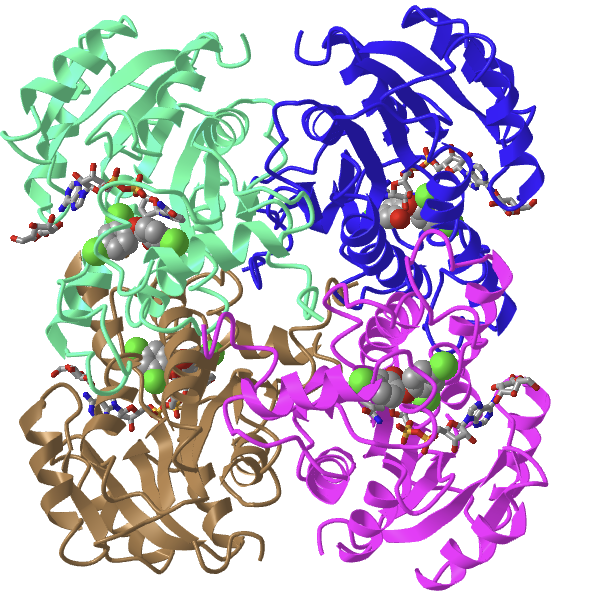
 Figure \(\PageIndex{12}\): enoyl-acyl carrier protein reductase (ER) in complex with NAD+ and triclosan (1QSG) (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...Z2ZCQaTYabQWa9
Figure \(\PageIndex{12}\): enoyl-acyl carrier protein reductase (ER) in complex with NAD+ and triclosan (1QSG) (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...Z2ZCQaTYabQWa9
Triclosan is used in many commercial products, even soaps, as it has broad-spectrum antimicrobial activity. In a world with increasing antibiotic resistance, it is unwise to use triclosan in soaps since the evidence shows that the surfactant properties of soaps are sufficient to remove bacteria from the skin. Triclosan is an active site inhibitor of bacterial enoyl-reductases but in humans, it appears to be an allosteric inhibitor, since it is bound at a protein-protein interface and not the active site.
Structure of Yeast Fatty Acid Synthase I (FAS1)
Now you have enough background to explore the actual structure of fatty acid synthase. Animal FAS1 is an α2 homodimer, some bacterial FASs are α6 hexamers, while fungal FASs are α6β6 dodecamers. In both cases, the monomers are multi-domain proteins with each domain having a different catalytic activity. There is another type of fatty acid synthase II (FAS2), that consists of separate enzymes each with their own catalytic function. Type II FAS around found in plants, most bacteria, and in mitochondria.
We'll explore the "more interesting" Type I and start with yeast (Saccharomyces cerevisiae) FAS (6QL5), whose structure has been solved with a regulatory γ subunit that 2.8 angstrom 6QL5. Figure \(\PageIndex{13}\) shows an interactive iCn3D model of the fatty acid synthase complex with bound gamma subunit from Saccharomyces cerevisiae (6QL5) very long load time.
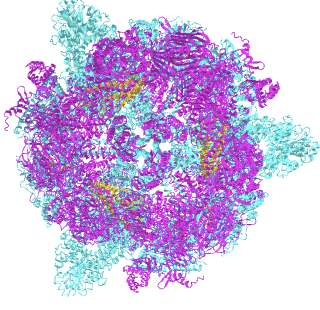
Figure \(\PageIndex{13}\): Fatty acid synthase complex with bound gamma subunit from Saccharomyces cerevisiae 6QL5. (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...YnMNSaTbTXYGe7 (very long load time).
The alpha subunits are shown in cyan and the beta subunits are shown in magenta. The regulatory γ subunit is shown in orange.
The 6 alpha subunits are arranged in an equatorial wheel or disk. The beta subunits are arranged in trimer top and bottom "domes" that cover the alpha subunit disk. Inside are 3 spaces or chambers where the reactions occur. 5 openings allow outside substrate access and six openings allow internal metabolite access. The FAS is more porous than in large structures with encapsulated volumes like the proteasome or viral capsids. It is more similar to pyruvate dehydrogenase. The ACP protein domain is found inside the structure and is tethered and "swingable" to allow the transfer internal transfer of acyl intermediates to the different enzymatic functional domains of the complex.
Figure \(\PageIndex{14}\) shows animated images of yeast fatty acid synthase complex, which should give you a sense of the arrangement of the equatorial alpha-helical wheel, the two beta subunit domes, and the interior and interior volumes available for reaction chemistry and intermediate movement. They switch between the full structure, followed by separate views of the alpha (cyan), beta (magenta), and regulatory (orange) chains.
| fatty acid synthase complex - side view | fatty acid synthase complex - top view |
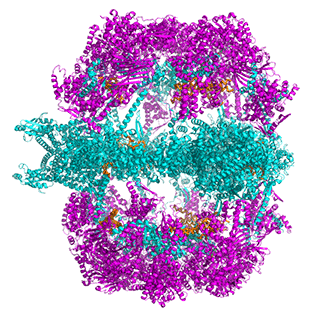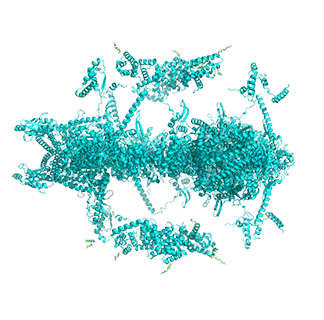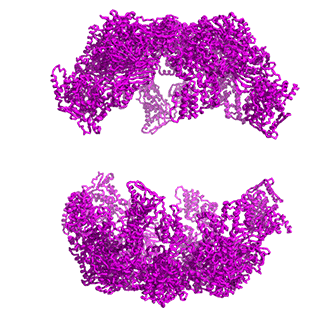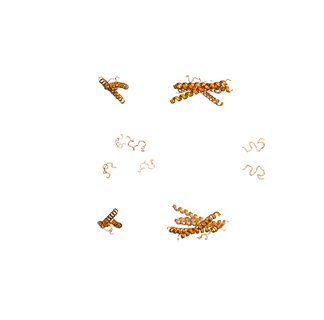 |
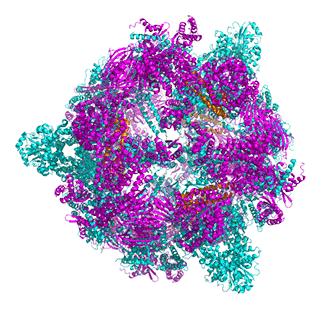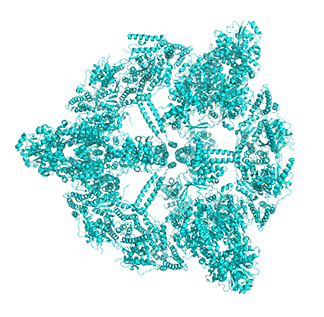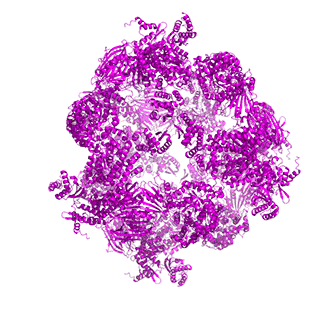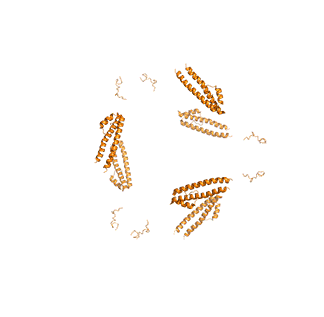 |
Figure \(\PageIndex{14}\): Alpha chains (cyan), beta chains (magenta), regulatory (orange)
The α and β monomers are multi-domain proteins with different domains having different catalytic properties. Just color coding the monomers in just one color doesn't give insight into the amazing catalytic properties of each chain. Figure \(\PageIndex{15}\) shows colored-coded domain and protein structures for yeast α6β6 dodecamer. The color-coding for the domains for each subunit shown at the bottom of panel A is also used for the domain structures in the actual protein structures.

Panel (A) shows the structure of S. cerevisiae FAS (3hmj). The side (left) and top view (right) with two β-subunits and two α-subunits are shown. Note that ACP is in the FAS interior, but is not shown. The MPT fold is comprised of both subunits and shown in cartoon representation (β part in brown and its α part in red). Nomenclature: acetyltransferase (AT), enoyl reductase (ER), dehydratase (DH), malonyl-palmitoyl-transferase (MPT), an acyl carrier protein (ACP), ketoacyl reductase (KR), ketoacyl synthase (KS) and phosphopantetheine transferase domain (PPT). Insertion elements are highlighted in grey; trimerization module (TM), 6-stranded β-sheet (SBS), hotdog-domain 2 (HDD2), dimerization module 1–4 (DM1-4), 4-helical bundle (4HB)). Please note that DM2 is not visible in this structure.
Panel(B) shows three yeast FAS barrels as a central D3-symmetric α hexamer (α6-wheel) and two C3-symmetric β trimers (β3-domes). β3-domes have been shifted for clarity (see arrows). ACP domains are shown for two α-subunits and are modeled by spheres in magenta. ACP linkers are indicated by dashed lines.
Panel (C) shows the post-translational modification of ACP. For phosphopantetheinylation, ACP and PPT have to physically interact. Fischer, M., Joppe, M., Mulinacci, B. et al. Analysis of the co-translational assembly of the fungal fatty acid synthase (FAS). Sci Rep 10, 895 (2020). https://doi.org/10.1038/s41598-020-57418-8. Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/
In Figure \(\PageIndex{16}\) we connect the domain structures of each monomer to the actual catalytic cycle of yeast FAS.
 |
|
Figure \(\PageIndex{16}\): Domain structure and the catalytic cycle of yeast fatty acid synthase (adapted from Singh et al. Cell, 180 (2020), https://doi.org/10.1016/j.cell.2020.02.034 and Fischer et al. Sci Rep 10, 895 (2020). https://doi.org/10.1038/s41598-020-57418-8. Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/
Figure \(\PageIndex{17}\) shows the actual structure of the isolated α and β monomers. The color coding of the domains is the same as used above.
| Fatty acid synthase 1 (FAS1) beta subunit (FAS1) (6U5U G chain) | Fatty acid synthase 2 (FAS2) alpha subunit (2UV8A - P19097 AlphaFold) |
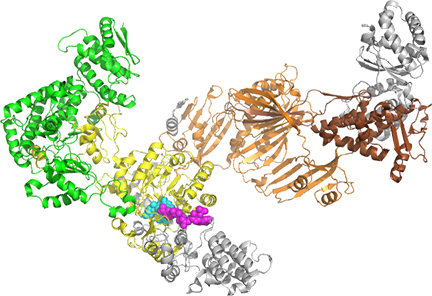 |
_alpha_subunit_(2UV8A%25C3%2582%25C2%25A0-%25C3%2582%25C2%25A0P19097%25C3%2582%25C2%25A0AlphaFold)V2.png?revision=1) |
Figure \(\PageIndex{17}\) shows the approximate domain structures and locations for the yeast fatty acid synthase 1 (FAS1) beta subunit (6U5U G chain) and fatty acid synthase 2 (FAS2) alpha subunit.
The color coding for the domains in each subunit is as follows: FAS1 beta subunit: AT, ER, DH, MPT, flavin mononucleotide (FMN) spacefill cyan, NADP nicotinamide-adenine dinucleotide phosphate (NAP), spacefill magenta; FAS2 alpha subunit: ACP, KR, KS, PPT
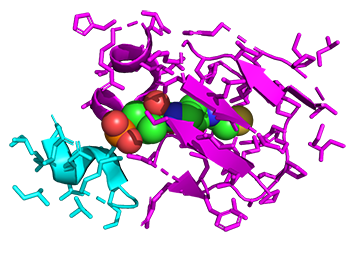
Now, let's zoom in to see the actual interactions of bound 4'-phosphopantetheine and flavin mononucleotide (FMN). Phosphopantetheine is bound between the alpha (cyan) and beta (magenta) subunits as shown in Figure \(\PageIndex{18}\).
| 4'-phosphopantetheine between alpha (cyan) and beta (magenta) subunits | flavin mononucleotide (FMN) within beta (FAS1) subunit |
 |
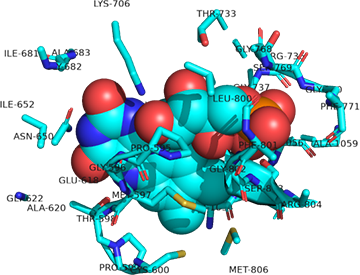 |
Figure \(\PageIndex{18}\):
The γ subunit crosses the entire inner cavity of FAS interfering with the activities of the reductases.
A central difference in the FAS1 found in humans and other animals occurs in the first reaction in which the acetyl group of acetyl-CoA is transferred to the pantetheine sulfhydryl of the acyl carrier protein (ACP) domain catalyzed by the malonyl‐/acetyltransferase (MAT) domain. It is then transferred to the active site cysteine of the β‐ketoacyl synthase (KS) domain. This allows the malonyl group to be transferred to the now free ACP domain for a second transfer reaction. A Claisen condensation reaction driven by the decarboxylation of the malonyl-ACP occurs to form the β‐ketoacyl intermediate bond to the KS domain. An alternative view of the mammalian fatty acid cycle is shown in Figure \(\PageIndex{19}\).

Panel (a) shows the priming of animal fatty acid synthesis. Panel (b) shows how in the first step, the substrate is selected by the MAT domain and transferred to the ACP domain (Step 2) from where it is passed on to the KS domain (Step 3). Important active site residues are highlighted and C161 is marked with an asterisk. The porcine FAS structure (2png) is shown. Domains of one protomer of FAS homodimer are colored. ACP, acyl carrier protein; DH, dehydratase; ER, enoyl reductase; FAS, fatty acid synthase; KR, ketoreductase; KS, ketosynthase; MAT, malonyl‐/acetyltransferase; PPant arm, expand fully in the figure; TE, thioesterase
Figure \(\PageIndex{20}\) shows the yeast FAS cycle (as a proxy for the mammalian cycle) placed in context with the mitochondrial matrix pathways that feed acetyl-Coa and NADPH into the fatty acid synthesis pathway.

Figure \(\PageIndex{20}\): yeast FAS cycle (as a proxy for the mammalian cycle) placed in context with the mitochondrial matrix pathways that acetyl-Coa and NADPH into fatty acid synthesis
Fatty acid Elongation and Desaturation
Overview
Elongation of fatty acids occurs in the cytoplasm of mammals from 16:0 made through fatty acid synthase. As with the synthesis of 16:0 by FAS, the elongation consists of two carbon addition driven by the decarboxylation of a malonyl-CoA substrate. Elongation is carried out by a family of ELOngation of Very Long-chain fatty acid enzymes (ELOVLs). Desaturation is catalyzed by Stearoyl-CoA Desaturases (SCD1 and SCD2) and Fatty Acid Desaturases (FADS1 and FADS2).
Figure \(\PageIndex{21}\) shows an overview of de novo fatty acid synthesis with elongation and desaturation in mammals as well as elongation and desaturation of dietary fatty acids.

FAs are elongated (ELOVL1–7) and/or desaturated (SCD, FADS) to generate complex FAs. Long-chain saturated FAs (LCSFA) and unsaturated FAs of ω9 and ω7 can be synthesized from palmitic acid (PA, C16:0) produced by the de novo FA synthesis. Long-chain unsaturated FAs of the ω6 and ω3 series can only be synthesized from essential diet-derived FAs (OA, oleic acid; NA, nervonic acid; ALA, α-linolenic acid; DHA, docosahexaenoic acids; LA, linoleic acid; ARA, arachidonic acid). SFA are saturated fatty acids.
Elongation
Mammals use endoplasmic reticulum very long-chain fatty acid enzymes (ELOVLs). There appear to be seven in mammals with different substrate specificities as shown in the figure above. The nomenclature for fatty acids is a bit strange. Those with 11-20 carbons are called long-chain FAs (LCFAs) and those with more than 20 carbons are very long-chain FAs (VLCFAs). Those with more than 26 carbons are called ultra long-chain FAs (ULCFAs), which are found in the skin, retina, meibomian gland, testis, and brain. The same four steps (condensation, reduction, dehydration, and reduction) used in FAS are used in elongation. The enzymes are abbreviated a bit differently (3-ketoacyl-CoA reductase or KAR, 3-hydroxyacyl-CoA dehydratase or HACD, trans-2-enoyl-CoA reductase or TER and LCFA). The enzyme is separate and not part of a multifunction complex like FAS. The elongation cycle is shown in Figure \(\PageIndex{22}\).
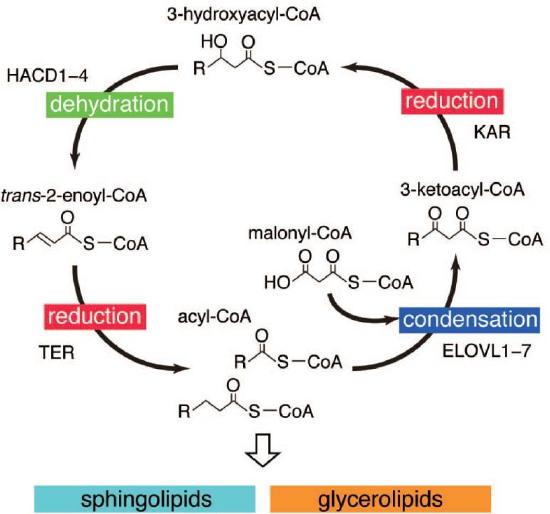
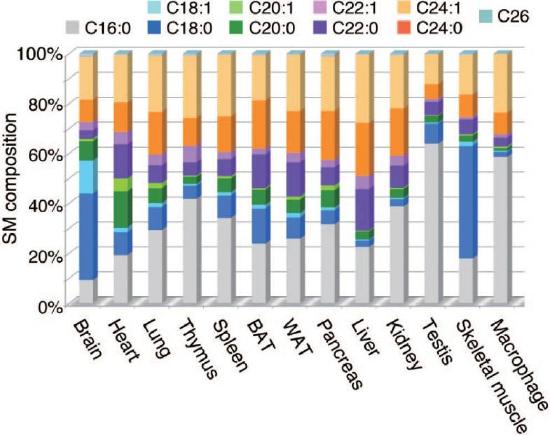
Figure \(\PageIndex{23}\) shows fatty acid composition in sphingomyelins which typically have long fatty acids.
Mechanism of ELOVL7
ELOVL condensing enzymes have a long hydrophobic tunnel and an active site nucleophilic histidine. More traditional nucleophiles like serine or cysteine were eliminated as potential candidates for the active site nucleophile since none were in proximity. The active site with a key HxxHH motif lies deep in the membrane. Nucleophilic catalysis leads to an N-acyl intermediate and is consistent with the observed ping-pong kinetics. An alternative reaction for a "bisubstrate" reaction is the formation of a ternary complex with both substrates. The narrow size of the binding pockets precludes this mechanism, so one product, after it is formed, must dissociate before the next substrate can bind. ELOVL7 preferentially catalyzes the elongation of C18-CoA presumably arising from the size of the acyl-binding product.
A plausible mechanism for human long-chain fatty acid enzyme 7 (ELOVL7) elongation is shown in Figure \(\PageIndex{24}\) and Figure \(\PageIndex{25}\). Figure \(\PageIndex{24}\) shows the beginning acylation step of the reaction.

Figure \(\PageIndex{24}\): Acylation step in the human long-chain fatty acid enzyme 7 (ELOVL7) elongation reaction
Figure \(\PageIndex{25}\) shows the condensation (with the elimination of CO2), followed by the reduction and dehydration of human ELOVL7.

Figure \(\PageIndex{26}\) shows an interactive iCn3D model of the human ELOVL fatty acid elongase 7 (ELOVL7) with bound 3-keto eicosanoyl CoA (6Y7F).
%25C2%25A0with_bound_3-keto_eicosanoyl_CoA_(6Y7F).png?revision=1&size=bestfit&width=390&height=310)
Figure \(\PageIndex{26}\): Human ELOVL fatty acid elongase 7 (ELOVL7) with bound 3-keto eicosanoyl CoA (6Y7F). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...eajMeiRgfHd9s9
The bound 3-keto eicosanoyl CoA is shown in spacefill, CPK colors. Key amino acids involved in catalysis are shown in sticks, CPK colors, and labeled.
Desaturation
The introduction of double bonds requires a different set of enzymes, acyl-coenzyme A (CoA) desaturases. Mammals have desaturases that can produce double bonds at Δ9, Δ6, and Δ5. There are two types, stearoyl-CoA desaturases (SCDs), which introduce a double bond at C9 of saturated fatty acid, and fatty acid desaturases (FADS), which work on unsaturated fatty acids. These are shown in Figure \(\PageIndex{20}\). Plants, but not mammals, have Δ12 and Δ15-desaturases, so they, but not mammals, can synthesize ω6 and ω3 fatty acids. These must be supplied by the diet as essential fatty acids and are precursors for the synthesis of longer fatty acids like arachidonic acid (20:4Δ5,8,11,14), necessary for prostaglandin synthesis (see below) and docosahexaenoic acids (DHA). A plausible mechanism for the stearoyl-CoA desaturase from castor seeds is shown below in Figures 27-30. The mechanisms and explanations are adapted from https://www.ebi.ac.uk/thornton-srv/m-csa/entry/136/. Creative Commons Attribution 4.0 International (CC BY 4.0) License

At the start of the reaction, ferredoxin, a small iron-sulfur protein donates a single electron, through Trp62, Asp228, and His146, to one of the Fe(III) centers in the desaturase. Ferredoxin then donates a second single electron, through Trp62, Asp228, and His146, to the second Fe(III) center. Both of these are shown in Figure \(\PageIndex{27}\)
Part 2 of the reaction is shown in Figure \(\PageIndex{28}\).

Figure \(\PageIndex{28}\): Part 2 of the stearoyl-CoA desaturase
The reduced charges on the Fe ions lead to loss of the oxide bridging ion (O2-) and its replacement with dioxygen. Water coordinates to one of the Fe(II)s, which causes it to donate a single electron to the dioxide molecule. This starts the first of two homolytic additions of the dioxygen molecule to both Fe(II) centers and the second Fe(II) center also donates a single electron to the dioxygen bridge.
Part 3 of the stearoyl-CoA desaturaseFigure \(\PageIndex{29}\) is shown below.

Part 4 of the stearoyl-CoA desaturase is shown in Figure \(\PageIndex{30}\).

In this reaction the hydroxide on the first Fe(IV) center deprotonates the stearoyl-[acyl-carrier protein] substrate, initiating the elimination of a hydride ion, which attacks the second Fe(IV) bound hydroxide. The excess electrons are donated singly to both Fe(IV) centers, regenerating the enzyme.
Figure \(\PageIndex{31}\) shows an interactive iCn3D model of the Stearoyl-acyl-carrier protein desaturase from castor seeds (1afr)
.png?revision=1)
Figure \(\PageIndex{31}\): Stearoyl-acyl-carrier protein desaturase from castor seeds (1afr) (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...UonrwyWfcg9wU8
An electron transport chain carries an electron from ferredoxin to one of the iron centers via several residues. A second electron is then carried to the second iron center. Several redox reactions take place involving oxygen, water, and iron centers, which result in the deprotonation of the substrate and the formation of a double bond.
There are two possible paths for the electron transport chain - the one described here has been chosen because of its analogy to what has been suggested for E. coli ribonucleotide reductase protein R2.
Mitochondrial FA Synthesis
Mitochondria also engage in limited fatty acid synthesis. They use separate discrete enzymes (at least 6) as do most bacteria. They don't employ a multienzyme complex as described above for yeast and mammals. Nevertheless, there are commonalities. Synthesis proceeds by the addition of two carbon units to a growing acyl-ACP chain using malonyl-CoA and the release of CO2 to drive the condensation process (in addition of course to ATP cleavage). The cytoplasmic fatty acid synthesis produces 16:0 (palmitate). In contrast, mitochondrial FAS produces two major products, 8:0 (octanoate). This is then converted to the 8C derivative lipoic acid. Their structures are shown in Figure \(\PageIndex{32}\).

We have seen lipoic acid before as a cofactor in pyruvate dehydrogenase and α-ketoglutarate dehydrogenase, two enzymes that catalyze the more difficult decarboxylation of α-ketoacids (which don't have a built-in electron sink at a β carbon adjacent to the departing carboxyl group. It is also used in branched-chain amino acid dehydrogenases. The absence of lipoic acid is lethal in mice.
Figure \(\PageIndex{33}\) shows the fatty acid synthesis pathway in mitochondria.
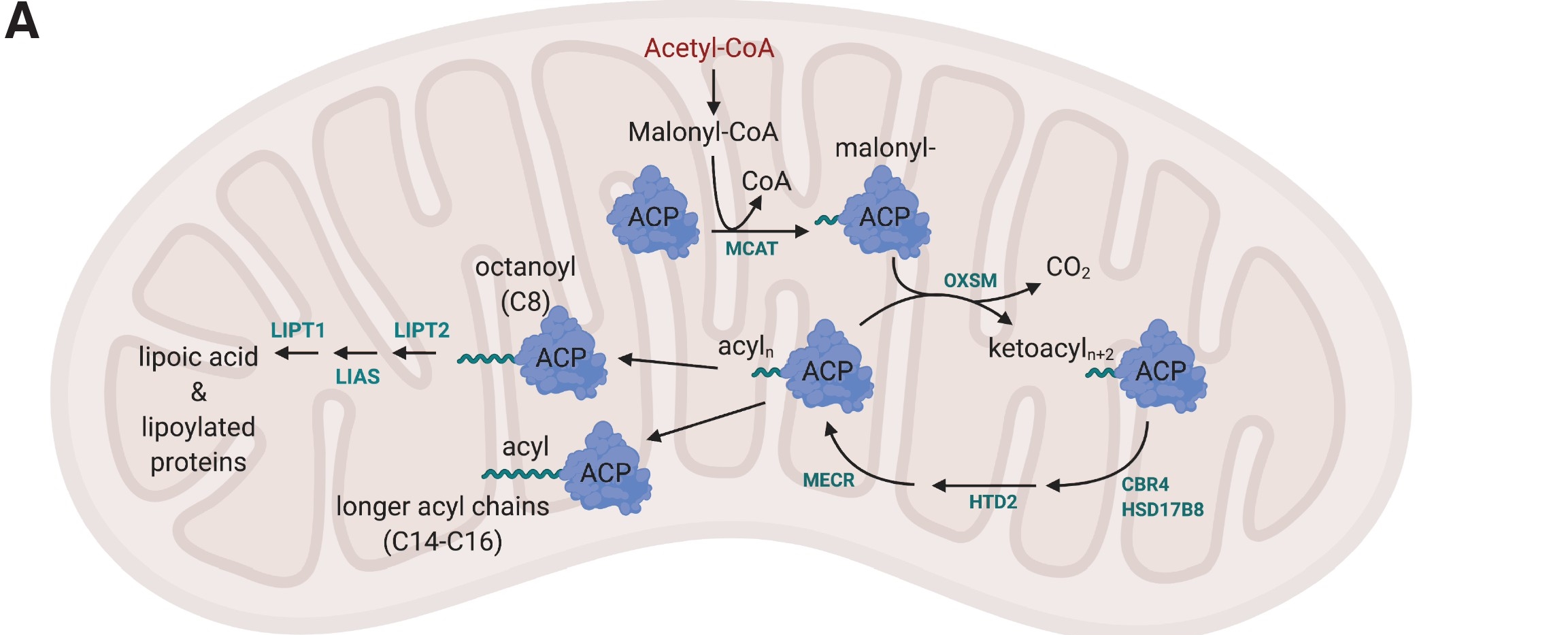
Figure \(\PageIndex{33}\): mitochondrial fatty acid synthesis pathway and downstream lipoic acid synthesis. Nowinski et al. eLife 2020;9:e58041. DOI: https://doi.org/10.7554/eLife.58041. Creative Commons Attribution License
Some of the enzymes in the pathway include the following: Mcat is malonyl-CoA ACP transacylase, and Oxsm is beta-ketoacyl synthase that catalyzes the Claisen condensation of malonyl-ACP with the growing acyl chain. CBR4 is 3-oxoacyl-[acyl-carrier-protein] reductase. HSD17B8 is (3R)-3-hydroxyacyl-CoA dehydrogenase. HTD2 is hydroxyacyl-thioester dehydratase type 2). Mecr is the terminal enoyl-[acyl-carrier-protein] reductase. LIPT2 is lipoyltransferase 2. LIAS is lipoyl synthase. LIPT1 (Lipoyltransferase 1) catalyzes the terminal step in lipoic acid and lipoylated protein synthesis.
It now appears that longer fatty acyl chains (up to 14C) are synthesized in mitochondria. Mutations in key enzymes above have been made that don't affect lipoic acid synthesis but impair mitochondrial function.
Eicosinoid and prostaglandin synthesis
As shown in Fig 20, arachidonic acid, 20:4Δ5,8,11,14, an ω-6 fatty acid (5,8,11,14- eicosatetraenoic acid), is synthesized from linoleic acid, 18:2Δ9,12, another ω-6 polyunsaturated fatty acid, through a series of elongation and desaturation steps. Arachidonic acid is released from membrane phospholipids on activation of phospholipase A2 and converted to a class of molecules called prostaglandins, which have powerful hormone-like effects.
The prostaglandins are part of a group of icosanoids derived from cleavage of membrane ω-3 (20:5Δ5,8,11,14,17, eicosapentaenoic acid) and ω-6 C20 fatty acids, which are metabolized to form leukotrienes (LTs), prostaglandins (PGs), prostacyclins (PCs), and thromboxanes (TXAs). These have differing and sometimes opposing physiologically effects in animals. Prostaglandins have been found in almost every tissue in humans and other animals
ω-3 and ω-3 and fatty acids
The most common polyunsaturated fats (PUFAs) in our diet are the ω-3 and ω-6 classes. Most abundant in the ω-6 class in plant food is linoleic acid (18:2 ω-6, or 18:2Δ9,12), while linolenic acid (18:3 ω-3 or 18:3Δ9,12,15) is the most abundant in the n-3ω class. These fatty acids are essential in that they are biological precursors for other PUFAs. Specifically,
- linoleic acid (18:2 ω-6, or 18:2Δ9,12) is a biosynthetic precursor of arachidonic acid (20:4 ω-6 or 20:4Δ5,8,11,14)
- linolenic acid (18:3 ω-3, or 18:3Δ9,12,15) is a biosynthetic precursor of eicosapentaenoic acid (EPA, 20:5 ω-3 or 20:5Δ5,8,11,14,17) and to a much smaller extent, docosahexaenoic acid (DHA, 22:6 ω-3 or 22:6Δ4,7,10,13,16,19).
These essential precursor fatty acids are substrates for intracellular enzymes such as elongases, desaturases, and beta-oxidation type enzymes in the endoplasmic reticulum and another organelle, the peroxisome (involved in the oxidative metabolism of straight chain and branched fatty acids, peroxide metabolism, and cholesterol/bile salt synthesis). Animals fed diets high in plant 18:2(n-6) fats accumulate 20:4(n-6) fatty acids in their tissues while those fed diets high in plant 18:3(n-3) accumulate 22:6(n-3). Animals fed diets high in fish oils accumulate 20:5 (EPA) and 22:6 (DHA) at the expense of 20:4(n-6).
Figure \(\PageIndex{34}\)

We will focus our attention on the conversion of arachidonic acid (20:4 ω-6 or 20:4Δ5,8,11,14) to prostaglandin G2 (PGG2) and H2 (PGH2) by the enzyme cyclooxygenase I and II. These enzymes are also called by their more formal names, prostaglandin endoperoxide H synthases (PGHSs) I and II. COX-1 prefers arachidonic acid as a substrate, while COX-2 has a broader substrate specificity and can use eicosapentaenoic acid (EPA, 20:5 ω-3 or 20:5Δ5,8,11,14,17) and even neutral substrate derivatives of arachidonic acid, including 2-arachidonoylglycerol and anandamide, both ligands for cannabinoid receptors.
COX 1/2 use two dioxygens and 2 electrons to make PGH2. Two different, separate but connected catalytic sites are used to catalyze the two steps (shown in Figure \(\PageIndex{34}\))
- Arachidonic acid (20:4Δ5,8,11,14) is converted to PGG2. This step is catalyzed by a bis-oxygenase domain. The word oxygenase is used since both atoms of dioxygen are added to the reactant. Bis indicates that two different molecules of dioxygen are added. Since one of the dioxygens added forms an endoperoxide cyclic bridge in PGG2, this enzymatic domain, and the entire enzyme is usually called cyclooxygenase (COX). The second dioxygen is added as a peroxide (both oxygen atoms of dioxygen added);
- Prostaglandin G2 (PGG2) is then converted to prostaglandin H2 (PGH2) in which the noncyclic peroxide form in the first step is converted to hydroxide. This is also a reduction step since the oxidation numbers of the oxygen atoms in the peroxide, -1, change to -2 (a gain of electrons) in the hydroxide in PGH2. This reaction is catalyzed by the peroxidase (POX) activity of the enzyme. A cofactor heme in the POX site facilitates catalysis.
The COX reaction proceeds through radical intermediates. but it requires the activation of peroxide by the heme in the POX site. We'll show two different mechanisms for both reactions (COX and POX) catalyzed by the enzyme prostaglandin endoperoxide H synthase, which we will also call COX.
The first mechanism shown below in Figure \(\PageIndex{35}\) emphasizes the free radical nature of the reactions. It shows just one amino acid (Tyr 385) involved in the reaction. Note that the first step in the reaction shows the added dioxygen as an excited state "singlet" O2, but it is parenthetically noted that it most likely involves O2 binding to and being activated by the heme iron in POX site.

A more detailed reaction mechanism for the synthesis of PGG2 by prostaglandin endoperoxide H synthase (COX), emphasizing the role of active site amino acid and the heme in the POX site, is shown in Figures 36-38 below. (The mechanism and its explanation are adapted from https://www.ebi.ac.uk/thornton-srv/m-csa/entry/37/. Creative Commons Attribution 4.0 International (CC BY 4.0) License)
The 1st step involves a peroxide (denoted H-O-O*) but it might also be a ROO peroxide or NO. The generation of a Tyr-385. free radical in the COX site. is critical for the reaction. This is preceded by the heme radical cation in the POX site.
Figure \(\PageIndex{36}\ below shows the first step in which His 207 deprotonates the alkyl peroxide, which then coordinates to the heme Fe(III) in the POX site.

In the next step, the peroxy bond donates two electrons to the heme Fe(III), one of which moves into the heme ring, forming Fe(IV). The heme ring then removes an electron from Tyr 385. This then removes a H. from the substrate, arachidonic acid. These steps are illustrated in Figure \(\PageIndex{37}\).

The heme site is not shown in the next steps. Next, the reactive dioxygen, itself a ground state diradical, form a bound to the carbon-free radical in arachidonic acid. The rest of the reaction ensues as shown in Figure \(\PageIndex{38}\).

Figure \(\PageIndex{39}\) shows an interactive iCn3D model of the mouse cyclooxygenase 2 (5COX)
.png?revision=1&size=bestfit&width=430)
Figure \(\PageIndex{39}\): mouse cyclooxygenase 2 (5COX) (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...wt9ECkgsyu4Vs8
Here is a model of human cyclooxygenase 1 structure (6Y3C): https://structure.ncbi.nlm.nih.gov/i...joK7oV63vLSi1A
Figure \(\PageIndex{39}\) shows an interactive iCn3D model of the mouse COX-2 with bound arachidonic acid (3KRK)
_.png?revision=1)
Figure \(\PageIndex{39}\): Mouse COX-2 with bound arachidonic acid (3KRK) (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...yAtJnTkTmuNreA
The following key binding and catalytic residues are shown and labeled:
-
Arg-120 and Tyr-355 are close to the carboxylate of AA
-
Phe205, Phe209, Val228, Val344, Phe381, and Leu534 form a hydrophobic groove for the ω-end of AA.
-
Ser 530, which is above this, gets acetylated by aspirin
-
Tyr 385, near C13 in arachidonic acid, forms the free radical which removes a single electron from C13
Prostaglandins, which were first isolated from prostate glands, serve as powerful, but labile local hormones which are mediators of pain, inflammation, and immune and clotting activity. The cyclooxygenase activity is inhibited by aspirin, which probably accounts for most of its anti-inflammatory and analgesic properties. Aspirin, acetylsalicylic acid, acetylates a reactive Ser 530 in the active site. Another nonsteroidal anti-inflammatory drug (NSAID) with similar properties is Ibuprofen (Advil). Acetaminophen (Tylenol) is also considered a member of this drug class, even though it doesn't have anti-inflammatory properties. The question has arisen as to why. It now turns out that there are three different types of COX, I, II, and III. COX III is expressed in the brain and might be involved in pain pathways. Acetaminophen appears to work on this COX, as shown in Table \(\PageIndex{1}\) below (Bazan et al.).
| COX | Expression | Function | Inhibitors |
|---|---|---|---|
| COX 1 | constitutively | organ pain, platelet function, stomach protection | NSAIDs including aspirin |
| COX 2 | induced by growth factors, neurotransmitters, inflammatory cytokines, oxidative stress, and injury. Constitutively in the brain, kidney | Inducible COX2: inflammation, pain, fever Constitutive COX2: synaptic plasticity |
NSAIDs, COX 2 inhibitors including celecoxib (Celebrex ) which has few GI problems associated with its use |
| COX 3 | constitutively, high in brain, heart | pain pathways, not inflammation pathways | acetaminophen (no GI problems, great fever reducer), some NSAIDs |
Fish n-3 fatty acids and health
We mentioned the importance of arachidonic acid in signal transduction in the lipid chapter. In addition, the importance of n-3 fatty acids to health was discussed as well. As mentioned above, arachidonic acid is cleaved from the C2 or sn-2 position of membrane phospholipids and modified by cyclooxygenase or lipoxygenase to form prostaglandins and leukotrienes, both potent local biological mediators. Linoleic acid and 22:6n-3 (DHA or docosahexaenoic acid) are also found in membrane phospholipids at the sn-2 position. What is the mechanism for the health-protective effects of n-3 fatty acids like DHA?
In human tissue, DHA, 22:6n-3 or 22:6Δ5,8,11,14,17,20 is the most abundant n-3 polyunsaturated fatty acids (PUFAs). Since it is synthesized from linolenic acid (as is EPA), a deficiency of linolenic acid in the diet will lead to lowered levels of 22:6n-3 in tissues, with ensuing health effects. Since these lipids are involved in membrane structure, signal transduction, and hormone synthesis, diverse effects of dietary n-3 PUFA deficiency will be observed. 50% of all fatty acids in the sn1 and sn2 position of membrane phospholipids of rod outer segments (in the retina) are 22:6(n-3). Cognitive dysfunctions (loss of memory, etc.) have been linked to decreased levels of 22:6(n-3) in the brain. This fatty acid binds to retinoid X receptors which then activate (through linked binding reactions) nuclear receptors, leading to alterations in gene transcription in the CNS.
In other tissues, 22:6(n-3) rarely exceeds 10% of membrane fatty acids, but this percentage can be increased in cells with increases in a precursor, 20:5(n-3). DHA might affect lipid rafts in the membrane, which would affect the movement of important membrane protein receptors (and associated proteins) in the membrane, altering cell response to environmental stimuli. DHA and EPA affect arachidonic acid conversion to prostaglandins and leukotrienes. EPA binds less tightly to cyclooxygenase I and is a poor substrate for the enzyme, both effects which inhibit the formation of prostaglandins and signaling processes mediated by them. This explains why n-3 fatty acids have anti-inflammatory effects.
In addition, n-3 fatty acids have noticeable effects on gene transcription, which remain as long as these fatty acids are present in high levels in the diet These and other fatty acids bind to fatty acid-activated transcription factors called PPARs (peroxisome proliferator receptors - alpha, beta and gamma 1 and 2). These receptors regulate, through alterations in gene expression, proteins involved in lipid metabolism. Other fatty acid-dependent transcription factors are known as well. PPARs bind 20:5(n-3) with a micromolar Kd and change the conformation of the protein to a form than can bind other proteins, ultimately altering gene expression.
| Organ(s) | Effect | Mechanism acts through |
|---|---|---|
| central nervous system | improve cognitive function | membrane composition; retinoic X receptor alpha |
| retina | improve acuity | membrane composition |
| immune | immunosuppressive; anti-inflammatory | membrane composition; rafts |
| cardiovascular | anti-arrhythmia; anti-clotting |
membrane composition; rafts; eicosanoids |
| serum lipids | lowers triglycerides (a risk factor for cardiovascular. disease) | peroxisome proliferator receptor alpha and gamma |
| liver | decrease lipid synthesis; increase fatty acid oxid. decrease VLDL synthesis |
sterol reg. element bind. protein; PPAR alpha PPAR alpha |
Recognition of Unsaturated Fatty Acids by Membrane G-protein-coupled receptors
The positive health effects of the n-3 (ω-3) fatty acids appear to involve those fatty acids acting as hormones that bind to a special membrane receptor called G protein-coupled receptor 120 (GPCR120). We will discuss GPCRs in greater detail in Chapter 28.2. When a hormone bounds to the membrane GPCR (a transmembrane protein with 7 membrane-spanning helices), the intracellular domain of the GPC alters how it interacts with a variety of binding partners inside the cell. The GPCRs are given that name since most interact with heterotrimeric G proteins inside the cell. These G proteins have a β, γ, and a variety of different α subunits (αq, αi, αs). Activation of different α subunits leads to different downstream effects as part of an elaborate signaling system in the cell. When the health-promoting polyunsaturated n-3 (ω-3) fatty bind to GPCR120, here are some of the beneficial health effects on metabolism, as outlined in Table \(\PageIndex{3}\) below.
| Protein involved in GPCR120 signaling | Downstream effect |
| Gαq |
Increases Ca2+ levels in the cell, which act as a second signal (messenger) |
| Gαi |
Increases insulin secretion |
| Gαs | Controls fat synthesis |
| non G protein GPCR kinase (GRK)/β-arrestin 2 | Inhibits the NLRP3 inflammasome and is ant-iinflammatory |
The data in the table above is summarized in Figure \(\PageIndex{40}\) below.

Figure \(\PageIndex{40}\): Schematic overview of G protein–and arrestin–mediated GPR120 signaling and related functions. Chunyou Mao et al. Science 380, eadd6220 (2023). https://www.science.org/doi/10.1126/science.add6220. With permission from the AAAS.
GPCR120 and differentiate fatty acids based on the number and position of the double bond in the acyl chain of the ligand. Even though saturated fatty acids can bind to GPCR 120, only the n-3 (ω-3) fatty acids elicit the health effects.
CryoEM structures show the fatty acids buried in an L-shaped conformation in the GPCR. The protein engages in noncovalent interactions, specifically π:π interactions, with the double bonds in acyl chain. These interactions, which depend on the number of double bonds in the n-3 (ω-3) fatty acid differentially biases the GPCR towards the promotion of the specific effects for each different fatty acid.
Figure \(\PageIndex{41}\) shows an interactive iCn3D model of the eicosapentaenoic acid bound GPR120-Gi complex (8ID9).
.png?revision=1&size=bestfit&width=283&height=385)
Figure \(\PageIndex{41}\): Eicosapentaenoic acid bound GPR120-Gi complex (8ID9). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...vgMGJRTr3emNj7
The color coding is as follows:
- GPCR120 - cyan (the membrane would be perpendicular across the 7 membrane helices) with the bound EPA
- Gβ - magenta
- Gγ - blue
- EPA -spacefill
Figure \(\PageIndex{42}\) shows an interactive iCn3D model showing the nonpolar environment of eicosapentaenoic acid bound to GPR120 (8ID9).
.png?revision=1&size=bestfit&width=381&height=491)
Figure \(\PageIndex{42}\): Nonpolar environment of eicosapentaenoic acid bound to GPR120 (8ID9). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...SszD1K3B9ieND7
Dark green shows the hydrophobic amino acids