
14.1: Regulation of Metabolic Pathways
- Page ID
- 15011


Exquisite mechanisms have evolved that control the flux of metabolites through metabolic pathways to insure that the output of the pathways meets biological demand and that energy in the form of ATP is not wasted by having opposing pathways run concomitantly in the same cell.
Enzymes can be regulated by changing the activity of a preexisting enzyme or changing the amount of an enzyme.
Changing the activity of a pre-existing enzyme
The quickest way to modulate the activity of an enzyme is to alter the activity of an enzyme that already exists in the cell. The list below, illustrated in the following figure, gives common ways to regulate enzyme activity
- Substrate availability: Substrates (reactants) bind to enzymes with a characteristic affinity (characterized by a dissociation constant) and a kinetic parameter called Km (units of molarity). If the actual concentration of a substrate in a cell is much less than the Km, the activity of the enzyme is very low. If the substrate concentration is much greater than Km, the enzyme's active site is saturated with substrate and the enzyme is maximally active.
- Product inhibition: A product of an enzyme-catalyzed reaction often resembles a starting reactant, so it should be clear that the product should also bind to the activity site, albeit probably with lower affinity. Under conditions in which the product of a reaction is present in high concentration, it would be energetically advantageous to the cell if no more product was synthesized. Product inhibition is hence commonly observed. Likewise, it is energetically advantageous to a cell if the end product of an entire pathway could likewise bind to the initial enzyme in the pathways and inhibit it, allowing the whole pathway to be inhibited. This type of feedback inhibition is commonly observed. Figure \(\PageIndex{1}\) shows product and end-product inhibition.
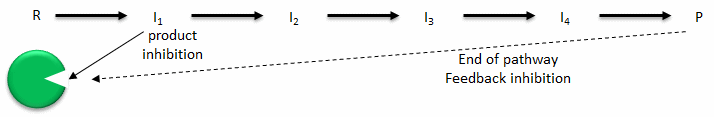
Figure \(\PageIndex{1}\): Product and end product inhibition of an enzyme
- Allosteric regulation: As many pathways are interconnected, it would be optimal if the molecules of one pathway affected the activity of enzymes in another interconnected pathway, even if the molecules in the first pathway are structurally dissimilar to reactants or products in a second pathway. Molecules that bind to sites on target enzymes other than the active site (allosteric sites) can regulate the activity of the target enzyme. These molecules can be structurally dissimilar to those that bind at the active site. They do so by conformational changes which can either activate or inhibit the target enzyme's activity.
- pH and enzyme conformation: Changes in pH that can accompany metabolic processes such as respiration (aerobic glycolysis for example) can alter the conformation of an enzyme and hence enzyme activity. The initial changes are covalent (change in the protonation state of the protein) which can lead to an alteration in the delicate balance of forces that affect protein structure.
- pH and active site protonation state: Changes in pH can affect the protonation state of key amino acid side chains in the active site of proteins without affecting the local or global conformation of the protein. Catalysis may be affected if the mechanism of catalysis involves an active site nucleophile (for example), that must be deprotonated for activity.
- Covalent modification: Many if not most proteins are subjected to post-translational modifications which can affect enzyme activity through local or global shape changes, by promoting or inhibiting binding interaction of substrates and allosteric regulators, and even by changing the location of the protein within the cell. Proteins may be phosphorylated, acetylated, methylated, sulfated, glycosylated, amidated, hydroxylated, prenylated, or myristoylated, often in a reversible fashion. Some of these modifications are reversible. Regulation by phosphorylation through the action of kinases, and dephosphorylations by phosphates are extremely common. Control of the phosphorylation state is mediated through signal transduction processes starting at the cell membrane, leading to the activation or inhibition of protein kinases and phosphatases within the cell.
Figure \(\PageIndex{2}\) shows ways to regulate the activity of pre-existing enzymes.
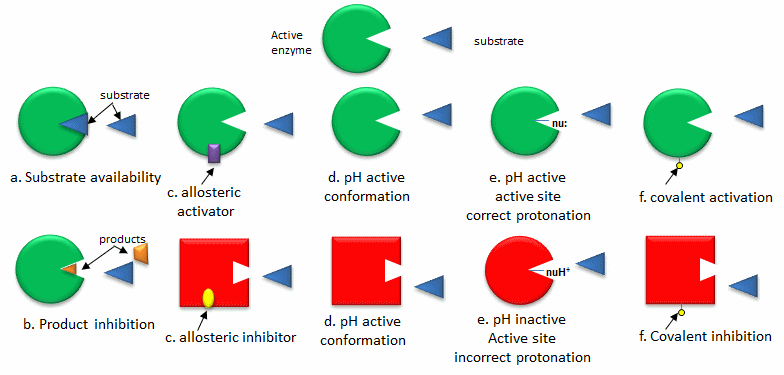
Figure \(\PageIndex{2}\): Ways to regulate the activity of pre-existing enzymes
Extracellular regulated kinase 2 (ERK2), also known as mitogen-activated protein kinase 2 (MAPK2) is a protein that plays a vital role in cell signaling across the cell membrane. Phosphorylation of ERK2 on Threonine 183 (Thr153) and Tyrosine 185 (Tyr185) leads to a structural change in the protein and the regulation of its activity.
Figure \(\PageIndex{3}\) shows an interactive iCn3D model showing the structural alignment of ERK2 in the dephosphorylated (5UMO) and phosphorylated (pY185) forms (2ERK). Toggle back and forth between the two structures with the "a" key.
_and_phospho_(pY185)_forms_(2ERK).png?revision=1&size=bestfit&width=407&height=252)
 Figure \(\PageIndex{3}\): Structural alignment of ERK2 in the dephosphorylated (5UMO) and phosphorylated (pY185) forms (2ERK). (Copyright; author via source). Click the image for a popup or use this external link:https://structure.ncbi.nlm.nih.gov/i...8iMQUVBbmMTRJ6
Figure \(\PageIndex{3}\): Structural alignment of ERK2 in the dephosphorylated (5UMO) and phosphorylated (pY185) forms (2ERK). (Copyright; author via source). Click the image for a popup or use this external link:https://structure.ncbi.nlm.nih.gov/i...8iMQUVBbmMTRJ6The residues that change significantly in conformation on phosphorylation are shown in blue. The side chain of tyrosine 185 in the unphosphorylated form is shown in CPK-colored sticks and labeled.
Regulation of single enzymes or entire pathways: Enzyme condensates
Single enzymes or all the enzymes of a given pathway can be coordinately regulated to maximize end-product output by organizing the enzymes in one large complex built from soluble enzymes that produce a "condensate" through a process similar to phase separation. Such condensates are shown for a series of enzymes in Figure \(\PageIndex{4}\).

Figure \(\PageIndex{4}\): Supramolecular assembly of enzyme condensates. Prouteau and Loewith. Biomolecules 2018, 8(4), 160; https://doi.org/10.3390/biom8040160. Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
The figure shows metabolism-related enzymes that form polymers in various organisms.
Panel (a) shows examples of metabolic enzymes observed to coalesce into cytosolic condensates.
Panel (b) top shows structures of metabolic enzymes that polymerize into filaments. The protomer of the polymer is shown above and placed into the filament below. These include P-Fructo-Kinase (4XYJ), cytidine triphosphate synthase (5U03), acetyl-CoA carboxylase (6G2D), glutamine synthetase (3FKY), mTORC1 (5FLC) from PDB files.
Panel (b) bottom shows the same structures from the Electron Microscopy Data Bank, including P-Fructo-Kinase filament (emd-8542), human CTP synthase filament (and-8474), human acetyl-CoA carboxylase with citrate (emd-4342), and the yeast glutamine synthetase filament.
Changing the amount of an enzyme
Another longer-duration method to modulate the activity of an enzyme is to alter the activity of an enzyme that already exists in the cell. Figure \(\PageIndex{5}\) shows ways in which enzyme concentrations are regulated.
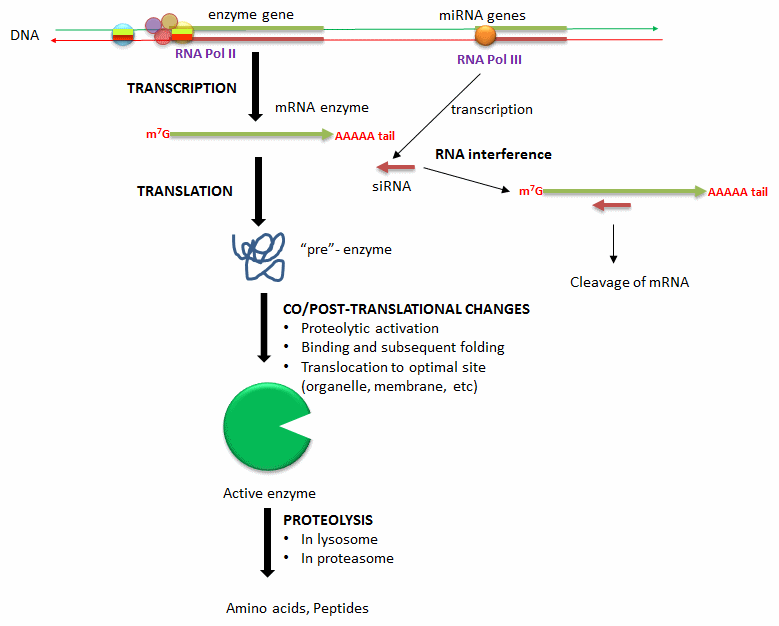
Methods include:
- Alterations in the transcription of enzyme's gene: Extracellular signals (hormones, neurotransmitters, etc) can lead to signal transduction responses and ultimate activation or inhibition of the transcription of the gene for a protein enzyme. These changes result from the recruitment of transcription factors (proteins) to DNA sequences that regulate the transcription of the enzyme gene.
- Degradation of messenger RNA for the enzyme: The levels of messenger RNA for a protein will directly determine the amount of that protein synthesized. Small inhibitor RNAs, derived from microRNA molecules transcribed from cellular DNA, can bind to specific sequences in the mRNA of a target enzyme. The resulting double-stranded RNA complex recruits an enzyme (Dicer) that cleaves the complex with the effect of decreasing the translation of the protein enzyme from its mRNA.
- Co/Post-translational changes: Once a protein enzyme is translated from its mRNA, it can undergo many changes that regulate its activity. Some proteins are synthesized in a "pre" form which must be cleaved in a targeted and limited fashion by proteases to activate the protein enzyme. Some proteins are not fully folded and must bind to other factors in the cell to adopt a catalytically active form. Finally, fully active protein can be fully proteolyzed by the proteasome, a complex within cells, or in lysosomes, which are organelles within cells containing proteolytic enzymes.
All proteins are ultimately regulated, if only by modulating the rates of their synthesis and degradation. However, some enzymes positioned at key points in metabolic pathways are ideal candidates for regulation, as their activity can affect the output of entire pathways. These enzymes typically have two common characteristics, they catalyze reactions far from equilibrium and they catalyze early committed steps in pathways.
Which Enzymes to Regulate: Reactions not at Equilibrium
The optimal enzymes for regulation are those at the beginning of pathways that carry out thermodynamically favored reactions. Why is the latter so important? These enzymes control the flux of metabolites through pathways, so to understand their regulation we can use the analogy of flow (or flux) of water from one container to another as illustrated in Figure \(\PageIndex{6}\).
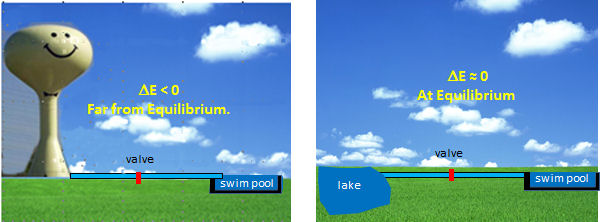
Figure \(\PageIndex{6}\): Regulation of water flow in pipe
Let's say you wish to fill a swimming pool at any desired height you wish and you have two ways to do so (see figure below). You could open a valve that controls the flow from your town's water tower to the pool. In this, the reaction (flow of water) is energetically (thermodynamically) favored given the difference in height of the water levels and the potential energy difference between the two. Even though flow (or flux) is cleared flavored, you can regulate it, from no flow to maximal flow, by opening and closing the valve (analogous to activating and inhibiting an enzyme). Your choices in the other scenario, filling the pool from a lake, are not so great. It would be hard to fill the water to the desired level (especially if it was an above-ground pool). It would be hard to regulate the flow.
By analogy, the best candidates for regulation are those enzymes whose reactions are thermodynamically favored (not at equilibrium) but which can be controlled by the mechanisms discussed in the previous section.
Which reactions are commonly not at equilibrium (i.e. ΔG<0 and usually also ΔG0 <0 if the ratio of products to reactants is not too high)? The answer is those that have reactants that are thermodynamically unstable compared to their reaction products. Several types of reactions often fit these criteria:
Hydrolysis (or similar reactions) of anhydride or analogous motifs: The figure below shows molecules with similar "anhydride" motifs and the ΔG0 for hydrolysis of the molecules. Those with more negative ΔG0 values can transfer their phosphate group to ADP to make ATP, which is necessary to drive unfavorable biological reactions. Metabolic reactions that involve hydrolysis (or other types of transfer reaction of these groups) usually proceed with a negative ΔG0 and ΔG, making them prime candidates for pathway regulation. Many textbooks label these types of molecules as having "high energy" bonds. This is confusing to many students as bonds between atoms lower the energy compare to when the atoms are not bonded. It takes energy to break the "high" energy phosphoanhydride covalent bond. What make hydrolysis of the molecules below so exergonic is that more energy is released on bond formation within the new products than was required to break the bonds in the reactants. In addition, other effects such as preferential hydration of the products, lower charge density in the products, and less competing resonances in the products all contribute to the thermodynamically favorable hydrolysis of the reactants.
Figure \(\PageIndex{7}\) shows thermodynamically unstable molecules (compared to their reaction products in aqueous solutions).

Thioesters (such as Acetyl-SCoA) are also included as they have the same negative ΔG0 of hydrolysis as ATP, even though they lack an "anhydride" motif. Thioesters are destabilized compared to their hydrolysis products and in comparison to esters made with alcohol since the C-S bond is weaker. Why?
Redox reactions: Everyone knows that redox reactions are thermodynamically favored if the oxidizing agent deployed is strong enough. The oxidation reactions of hydrocarbons, sugars, and fats by dioxygen are clearly exergonic (we do call these combustion reactions after all). What about redox reactions with less powerful oxidants? NAD+ is used frequently as a biological oxidizing agent. Are all these reactions as favored as combustion? Hardly so. Remember that in every redox reaction, an oxidizing and reducing agent react to form another oxidizing and reducing agent. Consider the following reaction:
Pyruvate + NADH ↔ Lactate + NAD+.
This reaction can go either way and is reversible. The above form is written in the favored direction in anaerobic metabolism when both Pyr and NADH levels are high. Although the ΔG0 favors the oxidation of lactate, given the high concentration of Pyr and NADH, the reaction is driven in the opposite direction and proceeds as shown. To determine if a redox reaction is favored and likely to occur (and possibly be regulated), the ΔG0 for a redox reaction should be calculated from standard reduction potentials, using the formula ΔG0 = -nFE0.
Which Enzymes to Regulate: Those catalyzing committed steps in pathways
The best enzymes to regulate are those that catalyze the first committed step in the reaction pathway. The committed step proceeds with a ΔG0 < 0 and is essentially irreversible. These reactions often occur from key metabolic intermediates that are immediately before or proximal to branches in reaction pathways. Two examples of key intermediates at branch points of metabolic pathways are shown in Figure \(\PageIndex{8}\) shows the reactions for the production and use of the intermediate glucose-6-phosphate.

Figure \(\PageIndex{8}\): Reactions for the production and use of the intermediate glucose-6-phosphate
Figure \(\PageIndex{10}\) shows reactions for the production and use of the intermediate acetyl-CoA.
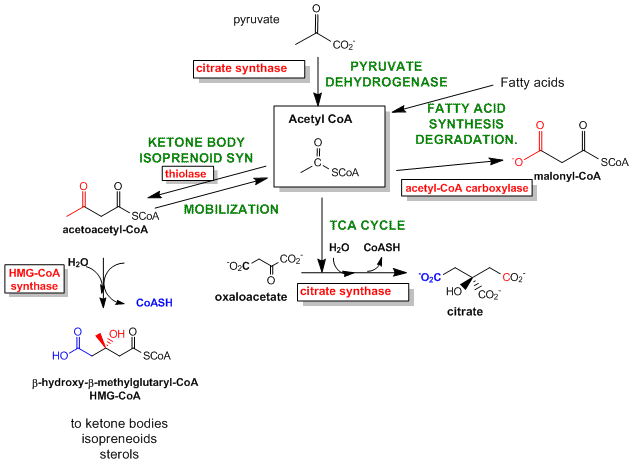
Figure \(\PageIndex{10}\): Reactions for the production and use of the intermediate acetyl-CoA
In reality, metabolic regulation is more complex and is distributed to many steps in a reaction pathway in ways that might not be evident without details mathematical analyses. We will discuss that in the next sections on metabolic control analysis.