
5.4: Regulation of Transcription
- Page ID
- 1653

The processes described above are required whenever any gene is transcribed. But what determines which genes are transcribed at a given time. What are the molecular switches that turn transcription on or off? Although there are entire books written on this one topic, the basic mechanism by which transcription is regulated depends on highly specific interactions between transcription regulating proteins and regulatory sequences on DNA.
We know that promoters indicate where transcription begins, but what determines that a given gene will be transcribed? In addition to the promoter sequences required for transcription initiation, genes have additional regulatory sequences (sequences of DNA on the same DNA molecule as the gene) that control when a gene is transcribed. Regulatory sequences are bound tightly and specifically by transcriptional regulators, proteins that can recognize DNA sequences and bind to them. The binding of such proteins to the DNA can regulate transcription by preventing or increasing transcription from a particular promoter.
Regulation in Prokaryotes
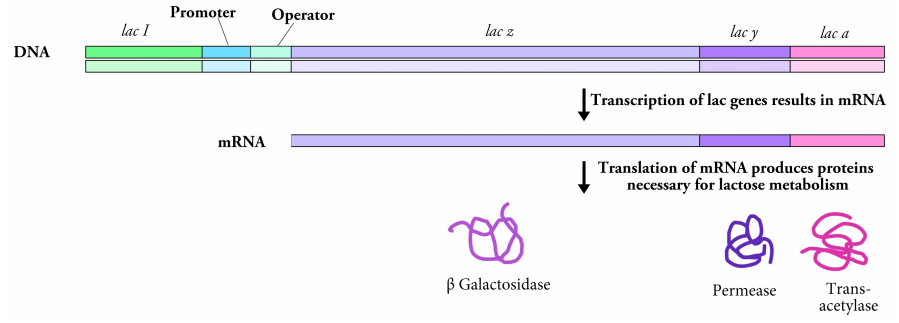
Let us first consider an example from prokaryotes. In bacteria, genes are often clustered in groups, such that genes that need to be expressed at the same time are next to each other and all of them are controlled as a single unit by the same promoter. The lac operon, shown in Figure 5.4.2, is one such group of genes that encode proteins needed for the uptake and breakdown of the sugar lactose. The three genes of the lac operon, lac z, lac y and lac a are controlled by a single promoter.
Bacterial cells generally prefer to use glucose for their energy needs, but if glucose is unavailable, and lactose is present, the bacteria will take up lactose and break it down for energy. Since the proteins for taking up and breaking down lactose are only needed when glucose is absent and lactose is available, the bacterial cells need a way to express the genes of the lac operon only under those conditions. At times when lactose is absent, the cells do not need to express these genes.
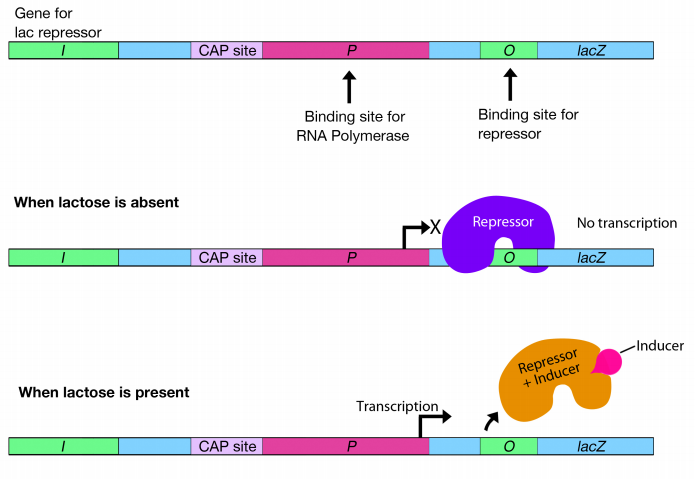
How do bacteria achieve this? Transcription of the lac cluster of genes is primarily controlled by a repressor protein that binds to a region of the DNA just downstream of the -10 sequence of the lac promoter. Recall that the promoter is where the RNA polymerase must bind to begin transcription. The place where the repressor is bound is called the operator (labeled O in the figure). When the repressor is bound at this position, it physically blocks the RNA polymerase from transcribing the genes, just as a vehicle blocking your driveway would prevent you from pulling out.
Obviously, if you want to leave, the vehicle that is blocking your path must be removed. Likewise, in order for transcription to occur, the repressor must be removed from the operator to clear the path for RNA polymerase. How is the repressor removed?
When the sugar lactose is present, it binds to the repressor, changing its conformation so that it no longer binds to the operator. When the repressor is no longer bound at the operator, the "road-block" in front of the RNA polymerase is removed, permitting the transcription of the genes of the lac operon.
Because the binding of the lactose induces the expression of the genes in the lac operon, lactose is called an inducer. (Technically, the inducer is allolactose, a molecule made from lactose by the cell, but the principle is the same.)
What makes this an especially effective control system is that the genes of the lac operon encode proteins that break down lactose. Turning on these genes requires lactose to be present. Once the lactose is broken down, the repressor binds to the operator once more and the lac genes are no longer expressed. This allows the genes to be expressed only when they are needed.
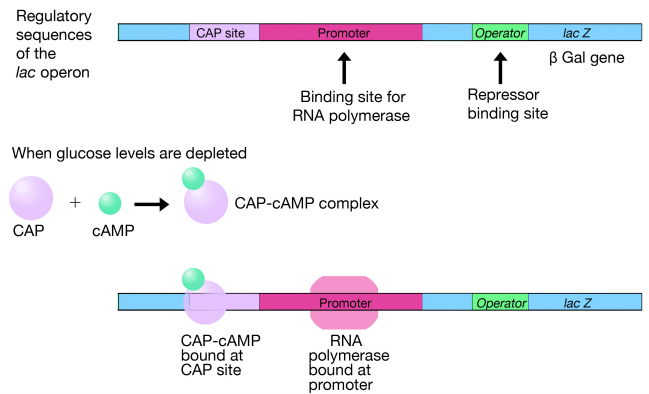
But how do glucose levels affect the expression of the lac genes? We noted earlier that if glucose was present, lactose would not be used. A second level of control is exerted by a protein called CAP that binds to a site adjacent to the promoter and recruits RNA polymerase to bind the lac promoter. When glucose is depleted, there is an increase in levels of cAMP which binds to CAP. The CAP cAMP complex then binds the CAP site, as shown in Figure 5.4.3. The combination of CAP binding and the lac repressor dissociating from the operator when lactose levels are high ensures high levels of transcription of the lac operon just when it is most needed. The CAP protein binding may be thought of as a green light for the RNA polymerase, while the removal of repressor is like the lifting of a barricade in front of it. When both conditions are met, the RNA polymerase transcribes the downstream genes.
The lac operon we have just described is a set of genes that are expressed only under the specific conditions of glucose depletion and lactose availability. Other genes may be expressed unless a particular condition is met. An example of this is the trp operon in bacterial cells, which encodes enzymes necessary for the synthesis of the amino acid tryptophan. These genes are expressed at all times, except when tryptophan is available from the cell's surroundings. This means that these genes must be prevented from being expressed in the presence of tryptophan. This is achieved by having a repressor protein that will bind to the operator only in the presence of tryptophan.
Regulation in Eukaryotes
Transcription in eukaryotes is also regulated by the binding of proteins to specific DNA sequences, but with some differences from the simple schemes outlined above. For most eukaryotic genes, general transcription factors and RNA polymerase (i.e., the basal transcription complex) are necessary, but not sufficient, for high levels of transcription.
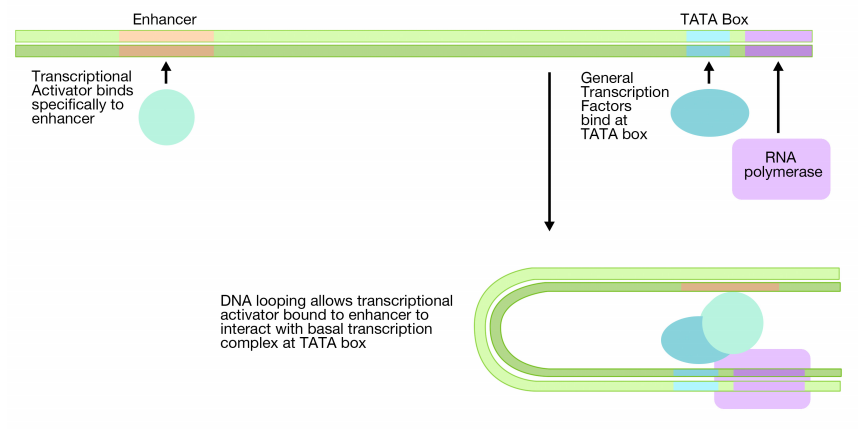
In eukaryotes, additional regulatory sequences called enhancers and the proteins that bind to the enhancers are needed to achieve high levels of transcription. Enhancers are DNA sequences that regulate the transcription of genes. Unlike prokaryotic regulatory sequences, enhancers don't need to be next to the gene they control. Often they are many kilobases away on the DNA. As the name suggests, enhancers can enhance (increase) transcription of a particular gene.
How can a DNA sequence far from the gene being transcribed affect the level of its transcription?
Enhancers work by binding proteins (transcriptional activators) that can, in turn, interact with the proteins bound at the promoter. The enhancer region of the DNA, with its associated transcriptional activator(s) can come in contact with the basal transcription complex that is bound at a distant TATA box by looping of the DNA (previous page). This allows the protein bound at the enhancer to make contact with the proteins in the basal transcription complex.
One way that the transcriptional activator bound to the enhancer increases the transcription from a distant promoter is that it increases the frequency and efficiency with which the basal transcription complex is formed at the promoter.
Another mechanism by which proteins bound at the enhancer can affect transcription is by recruiting to the promoter other proteins that can modify the structure of the chromatin in that region. As we noted earlier, in eukaryotes, DNA is packaged with proteins to form chromatin. When the DNA is tightly associated with these proteins, it is difficult to access for transcription. So proteins that can make the DNA more accessible to the transcription machinery can also play a role in the extent to which transcription occurs.
In addition to enhancers, there are also negative regulatory sequences called silencers. Such regulatory sequences bind to transcriptional repressor proteins. Transcriptional activators and repressors are modular proteins- they have a part that binds DNA and a part that activates or represses transcription by interacting with the basal transcription complex.